![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
|
Inheritance of acylsugar contents in tomatoes derived from an interspecific cross with the wild tomato Lycopersicon pennellii and their effect on spider mite repellence
ABSTRACT. Acylsugars present in Lycopersicon pennellii are responsible for the high levels of pest resistance often found in this wild tomato taxon. We investigated the inheritance of acylsugar contents in segregating populations of the interspecific tomato cross L. esculentum x L. pennellii and estimated correlations between leaflet acylsugar contents and the levels of mite repellence. Acylsugar contents were quantified with the Sommogy-Nelson colorimetric method in the acessions L. esculentum ‘TOM-584’ (P1, low acylsugars), L. pennellii ‘LA-716’ (P2, high acylsugars), in the interspecific F1 (P1 x P2) and in the F2 (P1 x P2) generations. Mite resistance was assessed by a repellence test. Broad-sense heritability of acylsugar contents was moderately high (h2b = 0.476). Frequency distributions in the P1, P2, F1 and F2 can be explained by the action of a single major locus, with near-complete dominance of the L. esculentum allele for low-acylsugar content over the L. pennellii allele for high content. Indirect selection for high levels of acylsugars in leaflets led to correlated increases in the levels of mite repellency, indicating that acylsugars may be the main factor involved in mite resistance. Key words: Tomato, Allelochemical, Acylsugars, Pest resistance, Trichomes, Spider mites, Tetranychus evansi, Lycopersicon esculentum, Lycopersicon pennellii
INTRODUCTION Tomatoes (Lycopersicon esculentum) are susceptible to a wide array of arthropod pests, particularly in tropical countries. Pest resistance levels of current cultivars are not sufficiently high to permit a significant reduction in the amount of pesticides used in the tomato crop. Development of cultivars with increased levels of arthropod resistance would be an important component for integrated pest management programs aimed at reducing the chemical sprays, allowing for diminished environmental impact (Barbosa, 1994). The wild tomato species Lycopersicon hirsutum var. hirsutum, L. hirsutum var. glabratum, L. peruvianum and L. pennellii are reported to be sources of resistance to many tomato pests (Gentile and Stoner, 1968; Gentile et al., 1968, 1969; Rick, 1973; Kennedy and Yamamoto, 1979; Williams et al., 1980; França et al., 1984a,b; Ecole et al., 1999). However, introgression of their arthropod resistance into the cultivated tomato is often limited by difficulty in maintaining the uniform infestations necessary to select for resistance (Stevens and Rick, 1986). Direct selection for pest resistance is expensive and slow, but indirect selection techniques based on correlated traits with high heritability could be used to speed up introgression (Juvik et al., 1982). Several allelochemicals present in wild Lycopersicon taxa have been associated with pest resistance: methyl-ketones such as 2-tridecanone in L. hirsutum var. glabratum (Williams et al., 1980; Fery and Kennedy, 1987; Weston et al., 1989; Eigenbrode and Trumble, 1993a,b; Barbosa, 1994; Maluf et al., 1997; Gonçalves et al., 1998); sesquiterpenes in L. hirsutum var. hirsutum (Snyder et al., 1987; Eigenbrode et al., 1994; Azevedo et al., 1999); acylsugars in L. pennellii (Goffreda et al., 1989). These compounds are found in glandular trichomes present in the leaf surface (Williams et al., 1980; Snyder and Carter, 1985; Carter and Snyder, 1985, 1986; Carter et al., 1989; Aragão, 1998), and are often associated with moderately high or high heritability values (Maluf et al., 1997; Freitas, 1999). Selection for high allelochemical content would therefore appear to be an efficient indirect selection technique for pest resistance (Juvik et al., 1982), as has been demonstrated with high 2-tridecanone or sesquiterpene (zingiberene) content and resistance to the South American tomato pinworm (Maluf et al., 1997; Azevedo et al., 1999), and spider mite repellence (Gonçalves et al., 1998; Campos, 1999). Spider mite (Tetranychus spp.) repellence mediated by these allelochemicals can be measured by quick, inexpensive techniques (Weston and Snyder, 1990), and can therefore be taken as indicative of the level of resistance to other arthropod pests (Aragão, 1998; Campos, 1999). The challenge posed by the silverleaf whitefly Bemisia argentifolii, also known as biotype B of the sweet potato whitefly B. tabaci, in the 1990’s, has brought renewed interest in breeding for pest resistance in tomatoes. This insect is now considered a major world pest of many horticultural and field crops. In the USA, damages have amounted to several billion dollars since it was reported for the first time in Florida greenhouses in 1986 (De Quattro et al., 1997). Schuster et al. (1989) estimated that Bemisia argentifolii caused 25 million dollars in damage to tomatoes in the State of Florida in 1989. Whitefly damages can be direct, when they feed on the sap and can cause irregular fruit ripening, or indirect, through the transmission of viral diseases, particularly geminiviruses (Van Lentere and Noldus, 1990; De Quattro et al., 1997). Lycopersicon pennellii accessions (particularly LA-716) have shown a very high level of resistance to the whitefly Bemisia tabaci/Bemisia argentifolii complex, as well as to aphids (Macrosiphum euphorbiae, Myzus persicae), mites and Lepidopteran pests (Gentile et al., 1968, 1969; Juvik et al., 1982; Goffreda et al., 1989), including the South American tomato pinworm Tuta (= Scrobipalpuloides) absoluta (França et al., 1989). Multiple pest resistance of L. pennellii ‘LA-716’ is due to the presence of type IV glandular trichomes that occupy the whole surface of the plant, together with the viscous phytochemicals that they secrete (Gentile et al., 1968; Goffreda et al., 1989). These phytochemicals are glucose and sucrose esters of fatty acids (acylsugars), and can play an important role in the resistance to tomato pests. The presence of type IV glandular trichomes that secrete acylsugars is controlled by at most two unlinked genes in crosses of L. pennellii with L. esculentum (Lenke and Mutschler, 1984). Purified acylsugars act by reducing feeding of the aphids Macrosiphum euphorbiae and Myzus persicae, by reducing feeding, larval development and survival of Helicoverpa zea and Spodoptera exigua, and by reducing oviposition and feeding of the leaf miner Liriomyza trifolii and of the silver leaf whitefly Bemisia argentifolii (Goffreda et al., 1988, 1989; Hawthorne et al., 1992; Rodriguez et al., 1993; Juvik et al., 1994; Liedl et al., 1995). While acylsugars are encountered mainly in L. pennellii and other wild species, their transfer to the cultivated tomato, L. esculentum, would contribute to increased levels of pest resistance in this commercial species. Information on the genetic control of acylsugar production and/or the chromosomal regions associated with that production would facilitate the transfer of these characteristics to the cultivated tomato. The present study reports on the inheritance of acylsugar contents in an interspecific cross of L. esculentum x L. pennellii. Also, plants with varying levels of acylsugars were selected from a segregating population from this interspecific cross, and tested for spider mite repellence (Weston and Snyder, 1990), in an attempt to verify whether indirect selection for high acylsugar contents would indeed lead to increased levels of arthropod resistance. MATERIAL AND METHODS Acylsugar determination Acylsugars in tomato leaflets were quantified by the method described by Resende (1999). Six leaf disks were taken from the upper third portion of each tomato plant with a 3/8" diameter cork borer (4.2-cm2 total leaf area), and placed in a test tube with 1 ml dichloromethane for extraction of acylsugars. The tubes were stirred with a Vortex mixer for 30 s. Leaflets were removed, the solvent was evaporated, and 0.5 ml 0.1 N sodium hydroxide dissolved in methanol (Merck) was added. The mixture was evaporated, the residue was maintained at 100oC and methanol was added three times at 2-min intervals to guarantee the completion of the saponification reaction. After evaporation of the methanol, the residue was dissolved in 0.4 ml water. Acylsugars can exist as both acylglucose and acylsucrose, therefore after saponification a mixture of glucose and sucrose is obtained. Sucrose was converted into glucose and fructose by adding 0.1 ml 0.04 N hydrochloric acid, boiling for 5 min and cooling. The Sommogy-Nelson reagent (Nelson, 1944) was added and the mixture was heated to boiling for 10 min and cooled to room temperature in a stream of cold water. Arsenomolibdate (0.5 ml) was added, the solution was stirred on a Vortex mixer for 15 s and the absorbance was measured at 540 nm in a spectrophotometer (Nelson, 1944). A standard curve was determined using standard glucose solutions, and absorbance readings were converted to concentrations expressed in nmol/cm2 of leaf area. Genetic material The tomato populations utilized in this studied were derived from an interspecific cross between L. esculentum ‘TOM-584’ and L. pennellii ‘LA-716’. ‘TOM-584’ is a tospovirus-resistant tomato breeding line with oblong-shaped fruit and indeterminate growth habit obtained from HortiAgro Sementes Ltda., Ijaci, MG, Brazil, with a genetic background similar to the standard open-pollinated Brazilian cultivar Santa Clara. ‘LA-716’ is an accession of L. pennellii obtained from the Tomato Genetics Stock Center/University of California, Davis, CA, USA, known to possess high levels of acylsugars in the leaves (Shapiro et al., 1994). F1 (TOM-584 x LA-716) seed was obtained by crossing TOM-584 (female parent) to LA-716 (pollen source). F1 plants were sib mated to obtain F2 (TOM-584 x LA-716) seed. Plants of TOM-584, LA-716, F1 (TOM-584 x LA-716) and F2 (TOM-584 x LA-716) were grown in a 128-cell tray spray filled with a commercial substrate, and transplanted 4 weeks later to 500-ml plastic pots filled a soil + humus + fertilizer potting mix. Total numbers of plants grown were 48, 48, 48 and 256, respectively, for TOM-584, LA-716, F1 (TOM-584 x LA-716) and F2 (TOM-584 x LA-716). Six to seven weeks after transplanting, leaf disks were collected as previously indicated from fully expanded leaflets of each plant, and used for acylsugar determinations. A block of six plants each of TOM-584, LA-716 and F1 (TOM-584 x LA-716) plus 32 plants of F2 (TOM-584 x LA-716) were sampled per day, so that all plants in the experiment were sampled within an eight-day period. Generation mean analysis and estimation of broad-sense heritability Mean acylsugar contents and their respective within-population variance estimates were obtained for TOM-584 (= P1), LA-716 (= P2), F1 (P1 x P2) and F2 (P1 x P2). Environmental variance (VE) was estimated as the geometric mean of the population variances of P1, P2 and F1. Genetic (VG) and phenotypic (VF) variances, as well as broad-sense heritability (h2b) were calculated as indicated by Mather and Jinks (1977). Generation mean analyses were performed on the P1, P2, F1 and F2 generations by the weighted least squares method (Mather and Jinks, 1977), to estimate the additive [d] and nonadditive [h] mean components and the mean degree of dominance (MDD = [h]/[d]). Frequency distribution of acylsugar content and test of the hypothesis of monogenic inheritance Frequency distributions of plant acylsugar contents were obtained for each generation studied in the trials. An arbitrary truncation point (TP) was chosen to permit a clear discrimination between the parental phenotypes: the TP value corresponded to an acylsugar content below which fell most of the plants of the resistant parent, and above which fell most of the plants of the susceptible parent. In this experiment TP was taken as equal to 40 nmol/cm2 leaf area, based on the observed frequency distributions in P1 and P2. Because of the continuous nature of the variable under study, classification of phenotypes into discrete classes (high/low acylsugars) was not used in this study. Data on the means and variances of P1 and P2 were used to estimate expected frequencies of plants below the TP in these populations, as well as in the F1 and F2, assuming monogenic inheritance and different presumed degrees of dominance. For each of the different presumed degrees of dominance, the monogenic hypothesis tested was based on the following assumptions and procedures: a) Acylsugar contents are normally distributed in populations P1, P2, F1 and F2. b) Parental generations P1 and P2 are assumed to have expected means and variances equal to their respective observed mean and variance values. Frequencies of plants where acylsugar content was below or equal to the truncation point (£TP) were obtained from the theoretical normal distribution assumed for P1 and P2, and were taken to represent the respective expected frequencies under the proposed model. c) The mean of the F1 population was admitted as being:
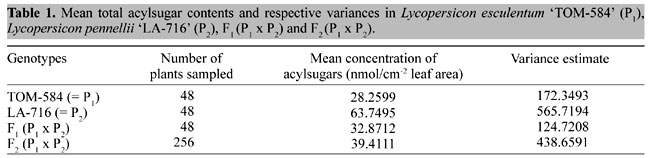
where: d) The frequency of F1 individuals with acylsugar values £TP was estimated from the normal distribution assumed for that population, and was assumed to be the true (expected) frequency under the model. e) under the hypothesis of monogenic inheritance, the expected frequency of plants in F2 with acylsugar values £TP was calculated as the weighted average of the expected frequencies in P1, P2 and F1 (weights 1:2:1, respectively), as estimated in (b) and (d). f) expected numbers of plants with acylsugar contents £TP in P1, P2, F1 and F2 were calculated by multiplying the expected frequencies (obtained in (b), (b), (d) and (e), respectively) by the total number of plants tested per generation. g) expected numbers of plants with values £TP in P1, P2 and F2 were compared with their respective observed values in each generation, and the significance of the deviations was estimated with a c2 test (with three degrees of freedom). h) significant c2 values would lead to the rejection of the hypothesis of monogenic inheritance under the degree of dominance considered. On the other hand, a nonsignificant value of c2 would lead to nonrejection of such a hypothesis. Values of c2 for each monogenic hypothesis/presumed MDD tested were plotted against their respective hypothetical MDD’s. The range of MDD values for which c2 values fall below the critical value would represent the MDD range for which the monogenic hypothesis cannot be rejected. Acylsugar content vs repellency to spider mites Plants were selected from population F2 (TOM-584 x LA-716) (hereafter designated BPX-370) based on contrasting acylsugar contents: 2 low acylsugar and 4 high acylsugar plants were selected of 256 F2 plants analyzed. These plant genotypes were subsequently maintained clonally via axillary shoot cuttings, and reserved for use in mite repellence tests. The six selected F2 genotypes were tested for mite repellence along with plants of TOM-584 (susceptible check), LA-716 (resistant check), and F1 (TOM-584 x LA-716) . Resistance to spider mites Tetranychus evansi was quantified through the thumbtack bioassay (Weston and Snyder, 1990). Spider mites, identified as Tetranychus evansi by the Department of Entomology, Universidade Federal de Lavras, Lavras, MG, Brazil, were collected from highly infested sweet potato, Ipomoea batatas, plants. Fully expanded tomato leaflets of similar sizes were removed from the apical region of flowering plants. One leaflet of each the 9 genotypes was attached to a sheet of styrofoam with a metallic thumbtack (9 mm in diameter) placed in the center of its adaxial surface. The 9 leaflets were randomly placed on the styrofoam sheet, and comprised one replication. Altogether, four replications were used. Ten female spider mites were transferred with a fine artist’s brush to the head of each thumbtack. The trial was carried out in a chamber at 16 ± 1ºC and 64 ± 4% relative humidity. Distances traveled by each mite onto the leaf surface were measured as the shortest distance between the mite and the thumbtack edge, and were recorded after 20, 40 and 60 min. Mites that stayed on the thumbtack were considered to have travelled a distance equal to zero. The contrast between the low acylsugar and the high acylsugar F2 genotypes was estimated. Also, the distances traveled by mites onto the selected F2 genotypes were regressed against their respective acylsugar contents. Data of TOM-584, LA-716 and the F1 were not included in the regression, but were analyzed separately through a standard analysis of variance and mean separation test, to confirm whether the thumbtack bioassay was able to detect differences in mite repellence between the check treatments. RESULTS AND DISCUSSION Acylsugar contents in L. pennellii ‘LA-716’ were twice as high as those found either in L. esculentum ‘TOM-584’ or in the F1 (TOM-584 x LA-716) (Table 1). F1 and F2 means were only slightly (ca. 14%) higher than those found in TOM-584, indicating that recessive gene(s) are responsible for the high acylsugar content found in LA-716.
Deviations from a simple additive-dominant model were not significant (Table 2), indicating that epistasis is not important in the expression of the trait. Both the additive ([a]) and the nonadditive ([d]) mean components differ significantly from zero, and the estimated mean degree of dominance was -0.74, an indication of near-complete dominance of the L. esculentum allele(s) over the L. pennelli allele(s). Broad-sense heritability estimated was moderately high (h2b = 0.476), indicating that nearly 50% of the total genetic variation can be explained by genetic effects (Table 2).
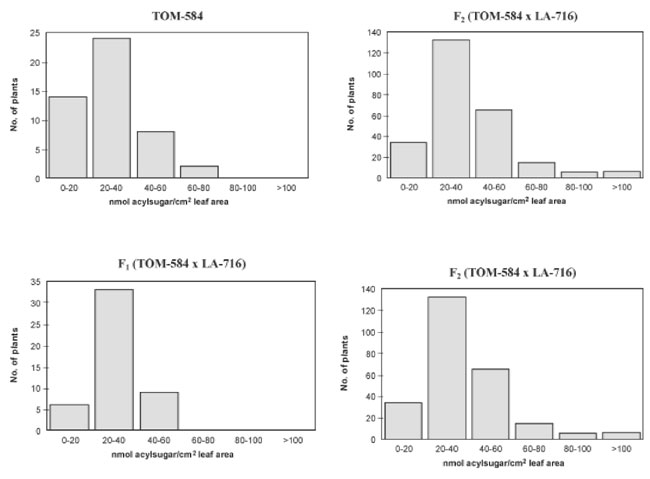
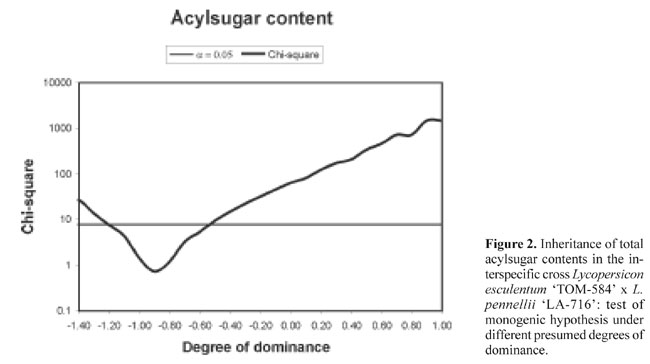
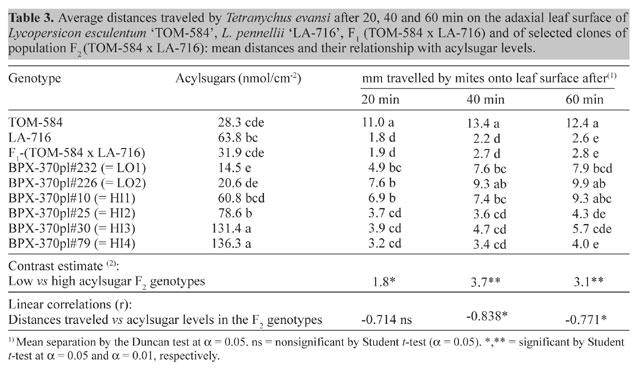
The environmental variance component (VE) includes both plant-to-plant and within plant variation. Because the within plant variation could probably be reduced by a more representative sampling of the leaflets, we presume that the estimated value for broad sense heritability could be increased. The frequency distributions of acylsugars among plants in TOM-584, LA-716, F1 and F2 indicate continuous variation for the trait, and no obvious discrete segregation pattern is indicated in the F2 (Figure 1). However, the hypothesis of monogenic inheritance is not rejected for the range of degrees of dominance between about -1.20 and -0.60 (Figure 2) - a range that encompasses the point estimate of -0.74 obtained in the generation mean analysis (Table 2). These distributions can therefore be explained by segregation of a single major locus with a degree of dominance within that range. Even though the action of modifier loci is not excluded, the action of a single major gene locus is evident, in which the recessive allele of L. pennellii is responsible for high acylsugar contents. Lenke and Mutschler (1984) indicated that type IV glandular trichomes, that exude acylsugars, show a rather simple inheritance: the number of type IV trichomes is under the control of two unlinked genes. There is no discrepancy, however, between their findings and those of our trial, because those authors did not measure acylsugars directly, but rather the morphological structures to which they are associated. On the other hand, their conclusion that inheritance of the number of type IV trichomes is not complex is compatible with our conclusions relative to acylsugar contents. In F2 genotypes selected for their contrasting acylsugar contents, spider mite Tetranychus evansi repellence was related to acylsugar concentration: distances travelled by mites onto the leaf surface after 20, 40 and 60 min were significantly shorter in the high acylsugar than in the low acylsugar genotypes (Table 3), and distances after 40 and 60 min were significantly and negatively correlated with acylsugar contents (Table 3). Increased mite repellence is an indirect response to selection for high acylsugars.
CONCLUSIONS It is apparent from the moderately high value of heritability and the rather simple inheritance pattern that acylsugar contents in plant leaves can effectively be increased by selection in segregating populations. Even though actual estimates of heritabilities for arthropod pest resistances in this interspecific cross are not available in the literature, it is expected that lower values of heritability might be obtained (Juvik et al., 1982), due to difficulties in ensuring uniform infestation in a host (plant) x pest x environment system. It is reasonable to assume that indirect selection for acylsugar contents (a higher heritability trait) can be more effective towards increasing arthropod resistance levels in tomato plants than direct selection for arthropod resistance per se. The mite repellence responses found in our experiment support this assertion. REFERENCES Aragão, C.A. (1998). Tricomas foliares associados à resistência ao ácaro rajado Tetranychus urticae Kach, em linhagens de tomateiro com alto teor de 2-tridicanona nos folíolos. M.S. thesis, Universidade Federal de Lavras, Lavras, MG, Brazil Azevedo, S.M., Maluf, W.R., Faria, M.V., Oliveira, A.C.B., Ribeiro, C.A., Gonçalves-Gervasio, R.C. and Santa-Cecília, L.V.C. (1999). Resistência à traça (Tuta absoluta) em genótipos de tomateiro com diferentes teores de sesquiterpenos nos folíolos. Hortic. Bras. 17: 273 (Abstract). Barbosa, L.V. (1994). Controle genético e mecanismo de resistência em Lycopersicon spp. à traça do tomateiro [Scrobipalpuloides absoluta (Meyrick, 1917) (Lepidoptera:Gelechiidae)]. M.S. thesis, Escola Superior de Agricultura de Lavras, Lavras, MG, Brazil. Campos, G.A. (1999). Inter-relações entre o teor de zingibereno, tipos de tricomas foliares e resistência a ácaros Tetranychus evansi em tomateiro. M.S. thesis, Universidade Federal de Lavras, Lavras, MG, Brazil. Carter, C.D. and Snyder, J.C. (1985). Mite responses in relation to trichomes of Lycopersicon esculentum x L. hirsutum F2 hibrids. Euphytica 34: 177-185. Carter, C.D. and Snyder, J.C. (1986). Mite responses and trichome characters in a full-sib F2 family of Lycopersicon esculentum x L. hirsutum. J. Am. Soc. Hortic. Sci. 111: 130-133. Carter, C.D., Sacalis, J.N. and Gianfagna, T.J. (1989). Zingiberene and resistance to Colorado Potato Beetle in Lycopersicon hirsutum f. hirsutum. J. Agric. Food Chem. 37: 206-210. De Quattro, J., Senft, D. and Wood, M. (1997). The whitefly plan - 5-year update. Agric. Res.: 4-12 (Feb. 1997). Ecole, C.C., Picanço, M., Jham, G.N. and Guedes, R.N.C. (1999). Variability in Lycopersicon hirsutum f. typicum and possible compounds involved in its resistance to Tuta absoluta. Agric. For. Entomol. 1: 249-254. Eigenbrode, S.D. and Trumble, J.T. (1993a). Antibiosis to beet armyworm (Spodoptera exigua) in Lycopersicon accessions. Hort. Sci. 28: 932-934. Eigenbrode, S.D. and Trumble, J.T. (1993b). Resistance to beet armyworm, Hemipterans, and Liriomyzia spp. in Lycopersicon accessions. J. Am. Soc. Hortic. Sci. 118: 525-530. Eigenbrode, S.D., Trumble, J.T., Millar, J.G. and White, K.K. (1994). Topical toxicity of tomato sesquiterpenes to the beet armyworm and the role of these compounds in resistance derived from an accession of Lycopersicon hirsutum f. typicum. J. Agric. Food Chem.42: 807-810. Fery, R.L. and Kennedy, G.G. (1987). Genetic analysis of 2-tridecanone concentration, leaf trichome characteristics and tobacco hornworm. resistance in tomato. J. Am. Soc. Hortic. Sci.112: 886-891. França, F.H., Maluf, W.R., Rossi, P.E.F. and Coelho, M.C.F. (1984a) Resistência em tomate à traça do tomateiro. In: Congresso Brasileiro de Entomologia, 9. Sociedade Entomológica do Brasil, Londrina, PR, Brazil, pp. 124. França, F.H., Maluf, W.R., Rossi, P.E.F., Miranda, J.E.C. and Coelho, M.C.F. (1984b). Avaliação e seleção em tomate visando resistência a traça do tomateiro. In: Congresso Brasileiro de Olericultura, 24. Resumos. Sociedade de Olericultura do Brasil, Jaboticabal, SP, Brazil, pp. 143. França, F.H., Maluf, W.R., Ferreira-Rossi, P.E., Miranda, J.E.C., Coelho, M.C.F., Castelo-Branco, M. and Resende, A.M. (1989). Breeding for resistance to Scrobipalpula absoluta (Meyrick) among Lycopersicon accessions in Brazil. In: Management Practices for Tomato and Pepper Production in the Tropics (Green, S.K., ed.). The Asian Vegetable Research and Development Center, Taiwan, China, 113-122. Freitas, J.A. (1999). Resistência genética de tomateiro Lycopersicon spp. à mosca branca Bemisia spp. mediada por zingibereno contido em tricomas glandulares. Ph.D. thesis, Universidade Federal de Lavras, Lavras, MG, Brazil. Gentile, A.G. and Stoner, A.K. (1968). Resistance in Lycopersicon species to the tobacco flea beetle. J. Econ. Entomol. 61: 1347-1349. Gentile, A.G., Webb, R. and Stoner, A.K. (1968). Resistance in Lycopersicon and Solanum to greenhouse whiteflies. J. Econ. Entomol. 61: 1355-1357. Gentile, A.G., Webb, R. and Stoner, A.K. (1969). Lycopersicon and Solanum resistant to the carmine and the two-spotted spider mite. J. Econ. Entomol. 62: 834-836. Goffreda, J.C., Mutschler, M.A. and Tingey, W.M. (1988). Feeding behavior of potato aphid affected by glandular trichomes of wild tomato. Entomol. Exp. Appl. 48: 101-107. Goffreda, J.C., Mutschler, M.A., Ave, D.A., Tingey, W.M. and Steffens, J.C. (1989). Aphid deterrence by glucose esters in glandular trichome exudate of wild tomato Lycopersicon pennellii. J. Chem. Ecol. 15: 2135-2147. Gonçalves, M.I.F., Maluf, W.R., Gomes, L.A.A. and Barbosa, L.V. (1998). Variation of 2-tridecanone level in tomato plant leaflets and resistance to two mite species (Tetranychus sp.). Euphytica 104: 33-38. Hawthorne, D.J., Shapiro, J.A., Tingey, W.M. and Mutschler, M.A. (1992). Trichome-borne and artificially applied acylsugars of wild tomato deter feeding and ovoposition of the leafminer Liriomyza trifolii. Entomol. Exp. Appl. 65: 65-73. Juvik, J.A., Berlinger, M.J., Ben-David, T. and Rudich, J. (1982). Resistance among accessions of genera Lycopersicon and Solanum to four of the main insect pests of tomato in Israel. Phytoparasitica 10: 145-156. Juvik, J.A., Shapiro, J.A., Young, T.E. and Mutschler, M.A. (1994). Acylglucose from wild tomato alters behavior and reduces growth and survival of Helicoverpa zea and Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol. 87: 482-492. Kennedy, G.G. and Yamamoto, R.T. (1979). A toxic factor causing resistance in a wild tomato to the tobacco hornworm and some other insects. Entomol. Exp. Appl. 26: 121-126. Lenke, C.A. and Mutschler, M.A. (1984). Inheritance of glandular trichomes in crosses between Lycopersicon esculentum and Lycopersicon pennellii. J. Am. Soc. Hortic. Sci. 109: 592-596. Liedl, B.E., Lawson, D.M., White, K.K., Shapiro, J.A., Cohen, D.E., Carson, W.G. and Mutschler, M.A. (1995). Acylsugar of wild tomato Lycopersicon pennelli alters settling and reduces oviposition of Bemisia argentifolii (Homoptera: Aleyrodidae). J. Econ. Entomol. 88: 742-748. Maluf, W.R., Barbosa, L.V. and Santa-Cecília, L.V.C. (1997). 2-Tridecanone-mediated mechanisms of resistance to the South American tomato pinworm Scrobipalpuloides absoluta (Meyrick, 1917) (Lepidoptera-Gelechiidae) in Lycopersicon spp. Euphytica 93: 189-194. Mather, K. and Jinks, J.L. (1997). Introduction to Biometrical Genetics. Cornell University Press, Ithaca, NY, USA. Nelson, N. (1944). A photometric adaptation of the Sommogy method for the determination of glucose. J. Biol. Chem. 153: 375-380. Resende, J.T.V. (1999). Teores de acilaçúcares mediadores da resistência a pragas e sua herança em folíolos de tomateiro, obtidos a partir do cruzamento interespecífico Lycopersicon esculentum x L. pennellii. M.S. thesis, Universidade Federal de Lavras, Lavras, MG, Brazil. Rick, C.M. (1973). Potential genetic resources in tomato species: clues from observations in native habitats. In: Genes, Enzymes and Populations (Srb, A.M., ed.). Plenum, New York, NY, USA, pp. 255-259. Rodriguez, A.E., Tingey, W.M. and Mutschler, M.A. (1993). Acylsugars of Lycopersicon pennelli deter settling and feeding of the green peach aphid (Homoptera: Aphididae). J. Econ. Entomol. 86: 34-49 Schuster, D.J., Everett, P.H., Price, J.F. and Krin, J.B. (1989). Suppression of the sweetpotato whitefly on commercial fresh market tomatoes. Proc. Fld. State Hortic. Soc. 102: 374-379. Shapiro, J.A., Steffens, J.C. and Mutschler, M.A. (1994). Acylsugars of the wild tomato Lycopersicon pennellii in relation to geographic distribution of the species. Biochem. Syst. Ecol. 22: 545-561. Snyder, J.C. and Carter, C.D. (1985). Trichomes on leaves of Lycopersicon hirsutum and L. esculentum and their hybrids. Euphytica 34: 53-64. Snyder, J.C., Johnson, D.A., Good, D.E. and Weston, P.A. (1987). Type VI trichome exudates from chemotypes of L. hirsutum and L. hirsutum f. glabratum. Rep. Tomato Genet. Coop. 37: 67-68. Stevens, M.A. and Rick, C.M. (1986). Genetics and breeding. In: The Tomato Crop: A Scientific Basis for Improvement (Atherton, J.G. and Rudich, J., eds.). Chapmann and Hall, New York, NY, USA, pp. 35-100. Van Lentere, J.C. and Noldus, L.P.J.J. (1990). Whitefly-plant relationships behavioural and ecological aspect. In: Whiteflies: Their Bionomics, Pest Status and Management (Gerling, D., ed.). Intercept, Hants, England, pp. 47-89. Weston, P.A. and Snyder, J.C. (1990). Thumbtack bioassay: a quick method of measuring plant resistance to twospotted spider mites (Acari: Tetranychidae). J. Econ. Entomol. 83: 501-504. Weston, P.A., Johnson, D.A., Burton, H.T. and Snyder, J.C. (1989). Trichome secretion composition, trichome densities, and spider mite resistance of ten accessions of Lycopersicon hirsutum. J. Am. Soc. Hortic. Sci. 114: 492-498. Williams, W.G., Kennedy, G.G., Yamamoto, E.T., Thacker, J.D. and Borner, J. (1980). 2-Tridecanone - a naturally occurring insecticide from the wild tomato Lycopersicon hirsutum f. glabratum. Science 207: 888-889. |