![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
|
Apomixis and cassava
ABSTRACT. Apomixis means seed formation without fertilization. In cassava (Manihot esculenta) it is an alternative to reproduction by cuttings, which normally transmits pathogens and leads to an accumulation of viral and bacterial diseases. Apomixis also assures preservation of heterosis and avoids genetic segregation. It occurs in wild relatives of cassava and has been transferred successfully from Manihot glaziovii and M. neusana. It is facultative, and occurs at a low frequency, ranging from 1-2%, and apparently is genetically different from apomixis in other crops. With selection, the frequency can reach 13%. Apomixis in cassava is frequently associated with aneuploidy but it does occur in some diploid types. It is due to the formation of aposporic sacs, which can easily be detected by clearing tissue preparations. Apomixis appears to have played an important role in speciation during the evolution of Manihot, since it leads to the maintenance and perpetuation of sterile interspecific hybridization. The use of apomixis in cassava breeding could lead to a boom in line improvement and commercial production. In addition to preserving superior genotypes, avoiding contamination of new plants, it would enable international programs to export their germplasm to destination countries. This would allow the use of superior genotypes even if apomixis occurs at a low frequency. A scheme to maximize benefits is to use diploid apomictic clones as maternal parents, which can be crossed with pollinators of polyploid interspecific hybrids, followed by selection among the progeny of new apomictic types that combine the heteroses of both interspecific hybridization and polyploidy. In addition, they acquire favored genes that have been transferred from the wild to the commercial crop. Key words: Apomixis, Triploid, Heterosis, Apospory, Interspecific hybridization
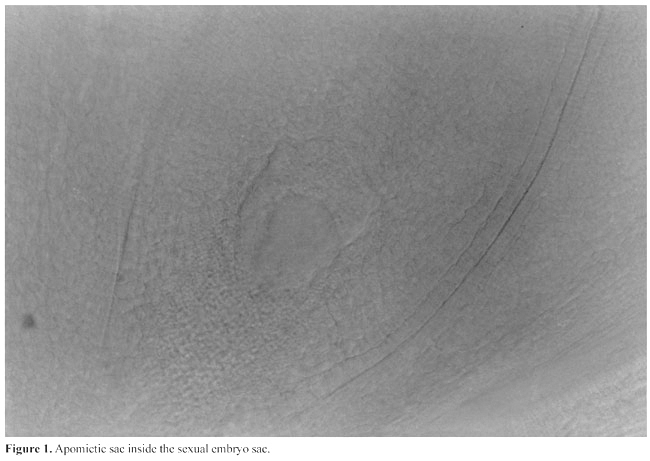
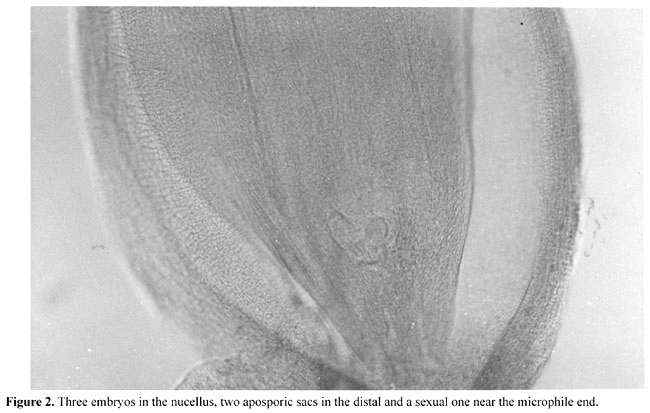
INTRODUCTION Apomixis means seed formation without fertilization. In cassava, it is an alternative to reproduction by cuttings, which normally is practiced by farmers. Propagation by apomixis avoids the transmission of viral and bacterial diseases that reduce productivity and could otherwise cause the extinction of superior genotypes. By using apomictic plants for propagation, systemic pathogens can be eliminated, and genetic segregation in the progeny is avoided. Seed produced through apomixis from a contaminated clone will be free from virus and bacteria and can begin a new cycle of clone life. If apomixis had been found in or introduced into the excellent Brazilian clones such as Guaxupe and Vassourinha, they would not have been become extinct, and could have been preserved forever. The use of apomixis for preserving superior genotypes and filtering microbial contamination would also benefit international cassava programs, which routinely export their germplasm. It would be sufficient in this case for the destination country to produce only one plant and further reproduce it vegetatively to be able to take advantage of the qualities of the new genotype. HISTORY OF APOMIXIS STUDY WITH CASSAVA Nassar and Ohair (1985) proposed the use of true seed to replace stem cuttings for cassava to eliminate the above-mentioned problems and potentially reduce production costs, and they emphasized the necessity of the production of easily germinating and apomictic clones. In the early 1980’s, we began selection for easily germinated seed of cassava clones. Apomictic lines were sought within the selected population. The criteria used were the transference of a group of characters through several generations from maternal plants to progeny, abundant fruit formation in plants that have sterile pollen, uniform progeny identical to the maternal type, and multiple seedlings per seed (Bashaw and Hanna, 1990). We found several distinct root, stem flower, and fruit characters, which we combined into groups of characters. Some of these characters were defined according to Rogers and Fleming (1973), while others such as flower color, fruit color, and root shape were defined according to Nassar and Grattapaglia (1986). For experimental confirmation of apomictic reproduction, controlled crosses were made between a clone with marker genes and the putative apomictic clones. A cytogenetic analysis of meiosis was also made. The offspring were analyzed for genetic segregation and by the criterion of transference of a maternal combination of morphological characters. One of the clones that showed strong indications of apomixis (the exhibition of a combination of characters in its progeny) was selected from the progeny of an interspecific hybrid with the variety Branca Santa Caterina. The interspecific hybrid was between Manihot dichotoma and cassava (Nassar, 1994, 1995). The second phase began in 1996, using a combination of embryonic, molecular and cytogenetic analyses for investigating apomixis in cassava. The molecular analysis was carried out with a RAPD assay. The rationale behind the analysis of putative apomixis with a RAPD assay is that a truly apomictic seedling would display a pattern of RAPD bands identical to the maternal parent, for all the markers surveyed. Polymorphism, i.e., non-maternal bands in the offspring or vice versa would reject the hypothesis of apomixis and indicate their zygotic nature. In addition to molecular techniques, putative apomictic clones had the morpho-structural development of embryo sacs studied histologically by clearing (Ogburia and Adachi, 1994). The transparent ovules were studied by differential interference contrast (DIC) optics. Both megasporangia and megagametophyte components were observed in large numbers of ovules (Nassar et al., 1998a,b). Pollen mother cells of the putative apomictic clones were also analyzed for meioses. The progenies of the clones had highly uniform DNA fingerprints. However, the markers readily showed that the plants were not derived from apomixis except for one individual in each progeny that had a pattern of RAPD bands identical to the maternal bands for all the primers. Aposporic sacs were found inside the sexual embryo sacs (Figure 1). Two embryo sacs were also found side by side in an ovule, which indicates the apospory nature of apomixis (Figure 2). The cytogenetic analysis showed that apomictic clones have 2n = 37, with a high rate of sterility, ranging from 20 to 30%. The second phase of the study further validated the occurrence of apomixis in cassava by presenting a combination of molecular and embryonic evidence from two putative apomictic clones. By analyzing a larger set of progeny, an advance over the earlier stage was made, making it possible to estimate about 2% facultative apomixis. Apomixis was found in a genotype derived from a new interspecific cross; parallel embryonic evidence was also generated, confirming the occurrence of apomixis and the aneuploid structure of the apomictic clones.
The third stage was to clarify the nature of apomixis in cassava and to study in detail the formation of aposporic embryo sacs. By clearing and using the DIC microscope we were able to photographically document the cassava embryo sac structure. The normal sacs had one egg, two polar nuclei and three antipodals. Synergids were occasionally seen. The egg was often inconspicuous. Antipodals were distinguished by a swollen teardrop shape, dense cytoplasm, chalazal position, and absence of a wall separating them from the sac cavity. In the hybrid of M. neusana Nassar with the cassava ovule it was possible to document photographically for the first time the presence of two sacs growing side by side (Nassar et al., 2000). One of these sacs apparently is the aposporous one while the other is the sexual embryo sac. The aposporous sacs lacked antipodes and had only one nucleus per sac. The development of two sacs jointly is a characteristic pattern of aposporic apomixis in cassava, and explains why we sometimes find two seedlings growing jointly from a single cassava seed. The two joint embryo sacs are frequently of the same size and phase of development. It seems that a certain external factor triggered growth at the moment of sexual embryo sac formation. Following megasporogenesis in ovules with aposporous embryo sacs, it was found that this process proceeds normally up to the point when the nucellar cell enlarges to form aposporous embryo sacs. In certain cases, these aposporous embryo sacs appear to develop faster than sexual embryo sacs, probably because they are not delayed by meiotic division. This is in accordance with Asker (1979) and Nogler (1984). In cases of development of aposporous embryo sacs from within the sexual cells, both the aposporous and the sexual embryo grew in parallel and finally coexisted. GENETIC AND CYTOGENETIC BASIS The controlled hybridization of the wild species M. neusana Nassar with cassava enabled the transfer of apomixis genes to the commercial species. While the hybrid (F1) did not acquire apomixis, as seen from the embryonic analysis, the second-generation plants did. This could mean that apomixis is controlled by more than one recessive gene, which act in an additive mode, probably carried on the same chromosome (Asker, 1979). In the cytogenetic study of an apomictic plant progeny 13 of 25 plants were sterile, and pollen viability ranged from 4 to 15%. Two plants had 2n + 1, while the rest were 2n. The other 12 plants were highly fertile with pollen viability ranging from 92 to 97%. Their chromosome number was 2n. The embryonic study revealed that all of the sterile plants were partially apomictic while the fertile plants were sexuals. Sterility apparently is associated with apomixis. Sterility is caused by consistent defects in meiosis due to a lack of pairing. All of these sterile plants showed asynapsis in meiotic metaphase. There were four to six univalents per cell. The irregular chromosome segregation in these sporocytes must have led to genetically unbalanced and aborted gametes. It seems that this sterility-causing genetic structure triggers certain apomixis genes to act. Apomixis will function and be established in such genotypes since it is favored by natural selection as it offers an escape from lethality, providing a perpetuation of the extant genotype. It can be concluded that: l) the nature of apomixis in cassava is different from other types found in other crops since it is present at very low levels, 1-2%. 2) It depends on meiotic irregularity, which often causes sterility in plants. 3) This sterility-causing genetic structure may trigger certain genes in cassava that activate a number of somatic cells in the nucellus or in the sexual embryo sac to form aposporic embryo sacs. 4) Natural selection would favor this apomictic genetic structure since it is an escape from extinction and a mode of perpetuation for the extant genotype. THE ROLE OF APOMIXIS IN MANIHOT SPECIATION Apomixis occurs frequently in Manihot species. We detected it in several species, namely M. neusana, M. glaziovii, M. cichotima and M. cearulescens. Combined with polyploidy and interspecific hybridization it may be responsible for rapid speciation in this genus. Polyploidy and interspecific hybridization offer the heterozygosity necessary for initial speciation. Sterile hybrids could be perpetuated through apomixis and they increase if favorable environmental conditions exist. Apomixis would enable the hybrids to maintain their heterogenic structure under these conditions, preserving precious genes for future recombination. The preserved diversity will be used when conditions change. In our living collection we noted several cases of truly breeding interspecific hybrids. Instead of segregation, the progeny were identical in morphology to the parents. This is the case for the interspecific hybrids between M. neusana and M. pohlii, M. neusana-M. cearulescens and M. neusana-M. pseudoglaziovii. Populations of these interspecific hybrids are being grown in our living collection and maintained through true seed. THE USE OF APOMIXIS IN CASSAVA BREEDING Recent studies have shown that cassava root productivity is due to a heterotic effect (Nassar, 1992; Rajendran et al., 2000). This heterosis can be preserved through apomixis. In the case of seed produced by international centers to be distributed in different countries, apomixis serves as an invaluable means to preserve superior genotypes. It will be possible to select superior plants from imported seeds. Those that are apomictic can be perpetuated vegetatively. This explains the superiority of the clones selected and released by IITA from Brazilian germplasm provided by this author in the 1970’s (Hahn, S.K., personal communication, 1982). The heterotic effect found in Brazilian germplasm is explained by this author as a result of hybridization between wild Manihot species and cultivated cassava. This hybridization not only leads to heterosis, but it also brings together genetic factors that may have the proper combinations for developing apomixis. Preserving heterosis through apomixis may be useful in another way. It can be used to manipulate a combination of both interspecific hybridization and polyploidy as follows: Interspecific hybrids normally exhibit high sterility. If the chromosome numbers were doubled by colchicine, fertility could be restored and further crosses would be possible. These polyploid interspecific hybrids can be crossed with partially apomictic plants (facultative with a low percentage of apomictic diploids) using the latter as females. Apomictic plants can be detected from the triploid seed produced and perpetuated vegetatively by the national programs that import them. ACKNOWLEDGMENTS Research supported partially by the Brazilian National Council of Research Development (CNPq). The living collection was established at the Universidade de Brasília with help of the Canadian International Development Research Center (IDRC), Ottawa, to whom this author is grateful. REFERENCES Asker, S. (1979). Progress in apomixis research. Hereditas 91: 231-240. Bashaw, E.C. and Hanna, W.W. (1990). Apomictic reproduction. In: Reproductive Versatility in the Grasses (Chapman, G.P., ed.). Cambridge University Press, Cambridge, pp. 100-130. Nassar, N.M.A. (1992). Cassava in South America: A plant breeder viewpoint. Cienc. Cult. (Braz. J. Ass. Adv. Sci.) 44: 25-28. Nassar, N.M.A. (1994). Selection of apomixis cassava clones. Cienc. Cult. (Braz. J. Assoc. Adv. Sci.) 48: 168-170. Nassar, N.M.A. (1995). Development and selection for apomixis in cassava Manihot esculenta Crantz. Can. J. Plant Sci. 74: 857-858. Nassar, N.M.A. and Grattapaglia, D. (1986). Variabilidade de clones de mandioca em relação a fertilidade e aspectos morfológicos. Turrialba 36: 555-559. Nassar, N.M.A. and Ohair, S.K. (1985). Variation among cassava clones in relation to seed germination. Indian J. Genet. 45: 394-398. Nassar, N.M.A.,Vieira M.A., Vieira, C. and Grattapaglia, D. (1998a). Evidence of apomixis in cassava, Manihot esculenta Crantz. Genet. Mol. Biol. 21: 527-530. Nassar, N.M.A., Vieira, M.A., Vieira, C. and Grattapaglia, D. (1998b). Molecular and embryonic evidence of apomixis in cassava interspecific hybrids (Manihot spp.). Can. J. Plant Sci. 78: 349-352. Nassar, N.M.A., Santos, E. and David, S. (2000). The transference of apomixis genes from Manihot neusana Nassar to cassava, M. esculenta Crantz. Hereditas 132: 167-170. Nogler, G.A. (1984). Gametophytic apomixis. In: Embryology of Angiosperms (Johri, B.M., ed.). Springer Verlag, Berlin, pp. 475-518. Ogburia, M.N. and Adachi, T. (1994). An improved cleared pistil technique for rapid in toto observation of embryo sac malformation in cassava, Manihot esculenta Crantz. Proceedings of the Second International Scientific Meeting on Cassava. Biotechnology Network, Indonesia, pp. 117-127. Rajendran, P.G., Ravindran, C.S., Nair, S.G. and Nayar, T.V.R. (2000). True Cassava Seeds (TCS) for Rapid Spread of the Crop in Non-Traditional Areas. Central Tuber Crops Research Institute Publication, Thiruvandram. Rogers, D.J. and Fleming, H.S. (1973). A monograph of Manihot esculenta Crantz with an explanation of taximetrics methods used. Econ. Bot. 27: 1-113. |