![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
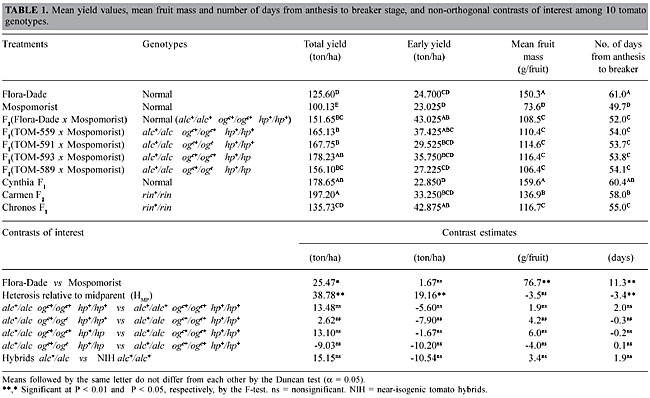
ABSTRACT. The effects of the fruit ripening mutant gene alcobaça (alc) and color development mutants, old gold-crimson (ogc) and high pigment (hp), on yield and post-harvest quality of tomato fruits were investigated. Five tomato hybrids were obtained by crossing near isogenic lines with Flora-Dade background [Flora-Dade (alc+/alc+ ogc+/ogc+ hp+/hp+), TOM-559 (alc/alc ogc+/ogc+ hp+/hp+), TOM-591 (alc/alc ogc/ogc hp+/hp+), TOM-593 (alc/alc ogc+/ogc+ hp/hp), and TOM-589 (alc/alc ogc/ogc hp/hp)] with the pollen parent line Mospomorist (alc+/alc+ ogc+/ogc+ hp+/hp+). Hybrid fruit was harvested at the breaker stage and stored on shelves at 15oC and 60% relative humidity for 16 days, and then evaluated for firmness, development of red color, and carotenoid contents. The different genotypic combinations at the loci alc, ogc and hp had no effect on fruit yield. The alc+/alc hybrid genotype significantly increased fruit firmness and significantly delayed the development of red color in maturing fruit. Simultaneous usage of ogc+/ogc and hp+/hp promoted an increase in the red color and lycopene content of alc+/alc hybrids, but did not have any additional effect on fruit firmness. Key words: Beta-carotene, Fruit firmness, Lycopene, Lycopersicon esculentum, Plant breeding, Ripening mutants INTRODUCTION The phenotypic and physiological effects of tomato ripening mutant genes have been studied in near-isogenic breeding lines. Some of these ripening mutants alter expression of one or more of the enzymes involved in fruit ripening or softening, and have been used in breeding programmes aimed at improving tomato fruit shelf life (Tigchelaar et al., 1978; Kopeliovitch et al., 1979). The mutant alcobaça (alc) is one of several ripening mutant loci that inhibit normal tomato ripening; it has been studied by tomato scientists in Brazil, as well as in other countries (Leal and Mizubuti, 1975; Lobo, 1981; Lobo et al., 1984; Mutschler, 1984a,b; Mutschler et al., 1992; Vilas Boas et al., 1999). The most evident effects of the homozygous genotype alc/alc are a marked increase in fruit firmness, a much-extended shelf life, and inhibition of carotenoid pigment synthesis; alc/alc fruit mature off-vine to a yellow color, which renders them unmarketable. Heterozygous alc+/alc hybrids are intermediate in firmness between their normal (alc+/alc+) and homozygous alcobaça (alc/alc) parents (Lobo, 1981; Mutschler et al., 1992; Freitas et al., 1998; Souza et al., 2001), indicative of partially dominant gene action (Freitas et al., 1998; Souza et al., 2001). Heterozygous alc+/alc fruit ripened off-vine mature to a nearly normal red color, without any apparent further deleterious effects (Leal and Mizubuti, 1975; Mutschler et al., 1992; Flori and Maluf, 1994; Resende et al., 1997; Freitas et al., 1998; Vilas Boas et al., 1999; Souza et al., 2001; Araújo et al., 2002). They are slower in ripening than normal alc+/alc+ genotypes, and therefore remain firm for a longer period of time; their rate of red color development during the ripening process is also slower than normal, but the fruit are still acceptable for marketing (Freitas et al., 1998). The alcobaça mutant can interact with ogc (old gold-crimson) and hp (high pigment), which are mutants that affect fruit carotenoid pigment contents (Araújo et al., 2002). These mutants increase fruit lycopene content, and therefore have a positive effect on fruit color and nutraceutical properties. The hp locus is a monogenically inherited mutation in chromosome 2 that increases fruit quality and the quantity of carotenoid pigments, especially lycopene and beta-carotene (Thompson et al., 1962). The old gold-crimson (ogc) locus is a monogenically inherited mutation, causing orange (old gold) colored petals and increased fruit lycopene content (Thompson et al., 1967). Epistatic effects among the loci alc, ogc and hp have been described (Araújo et al., 2002) in the inbred background Floradade, and specific homozygous or heterozygous combinations of ogc and/or hp were found to contribute to color improvement of alc+/alc genotypes. Some genotypic combinations reduced yield and fruit mass, but these could be background-specific, and might be overcome in hybrid combinations. We examined the effects of alc+/alc on yield and post-harvest quality of hybrid tomato fruit, and quantified the effects of heterozygotes at loci ogc and hp on fruit yield and quality traits of alc+/alc fruit, in order to determine the feasibility of the exploitation of these genotypic combinations for developing extended shelf life tomato hybrids. MATERIAL AND METHODS Five near-isogenic tomato lines in background Flora-Dade (Flora-Dade, TOM-559, TOM-591, TOM-593 and TOM-589), differing from each other at the loci alc, hp and ogc, were used as seed parents in combinations with the Mospomorist line (pollen source), resulting in five near-isogenic tomato hybrids (NIH). Flora-Dade is a standard open-pollinated cultivar (alc+/alc+ ogc+/ogc+ hp+/hp+) from the University of Florida/USA. TOM-559, TOM-591, TOM-593 and TOM-589 differ from Flora-Dade in being, respectively, alc/alc ogc+/ogc+ hp+/hp+ (alcobaça), alc/alc ogc/ogc hp+/hp+ (alcobaça, old gold-crimson), alc/alc ogc+/ogc+ hp/hp (alcobaça, high pigment) and alc/alc ogc/ogc hp/hp (alcobaça, old gold-crimson, high pigment). Mospomorist is a Moneymaker-type accession, obtained by the Institut National de Recherche Agronomique, Avignon, France, possessing multiple disease resistance and indeterminate growth habit. The five hybrids were tested along with the two open-pollinated alc+/alc+ ogc+/ogc+ hp+/hp+ lines (Flora-Dade, Mospomorist), two long shelf-life hybrids heterozygous at locus rin (rin+/rin) (Carmen F1, Chronos F1), and one normal (alc+/alc+ ogc+/ogc+ hp+/hp+) hybrid (Cynthia F1). The near-isogenic hybrids had the genotypic constitutions: F1(Flora-Dade x Mospomorist) = alc+/alc+ ogc+/ogc+ hp+/hp+= normal; F1(TOM-559 x Mospomorist) = (alc+/alc ogc+/ogc+ hp+/hp+) = heterozygous alcobaça; F1(TOM-591 x Mospomorist) = (alc+/alc ogc+/ogc hp+/hp+) = heterozygous alcobaça and crimson; F1(TOM-593 x Mospomorist) = (alc+/alc ogc+/ogc+ hp+/hp) = heterozygous alcobaça and high-pigment, and F1(TOM-589 x Mospomorist) = (alc+/alc ogc+/ogc hp+/hp) = heterozygous alcobaça and crimson and high-pigment. Except for Flora-Dade, all the genotypes had an indeterminate growth habit. The 10 genotypes were grown in a plastic greenhouse from July through December 1999, in a randomized complete block design, with four replications. Each plot was comprised of a single row with 11 plants. Plants were pruned to a single stem with five racemes. The plant density was 3.84 plants per square meter. Flowers were date-tagged at anthesis in order to determine the number of days between anthesis and the harvest of the fruits at the breaker stage. Multiple harvests (17) were made, spanning an 80-day period from late September through December 1999. Total yield (ton/ha), early yield (yield during the first 20 days of harvest), mean fruit mass (g/fruit), and the number of days from anthesis to breaker stage (harvest), were recorded. A sample of 12 fruits per plot was harvested at the breaker stage and, after being placed in a chamber at 15oC and 60% relative humidity, was evaluated for rate of loss of firmness, and evolution of red color during ripening. Fruit firmness (N/m2) was recorded at the equatorial surface for each individual fruit through a non-destructive flattening technique (Calbo and Calbo, 1989; Calbo and Nery, 1995). Firmness readings were taken at breaker stage (day 0) and at 2-day intervals until 16 days after harvest (d.a.h.). The same fruit samples were scored from 0 to 16 d.a.h. on a scale of 1 to 5, according to estimated percent colored surface of the fruit. Percent colored surface at day 0 (= breaker stage) was taken as score 1. Scores 2, 3, 4 and 5 corresponded to percent red colored fruit surface from 20 £ 40, 40 £ 60, 60 £ 80 and 80-100%, respectively. Non-linear regression models were fitted to the rate of firmness loss and to the rate of red color development, over time. Fruit firmness over time was fitted to an exponential decay curve, which was then used to estimate initial fruit firmness (= fruit firmness at the breaker stage), and the number of days necessary to reach a fruit firmness of 4 x 104 N/m2. A logistic Y = A/(1 + B*RX) model was fitted to fruit color score data over time (X = number of days after harvest; Y = fruit color score); scores for fruit color 12 d.a.h., and the mean number of days to reach color score 4.0 was then estimated from the adjusted curve. An additional sample of eight fruits per plot was used to quantify carotenoid pigments at three different stages of on-vine fruit ripening - breaker, intermediate and fully mature stage (corresponding to fruit color scores 1, 3 and 5, respectively). Epicarp-free longitudinal slices of fruit were taken from each eight-fruit sample, and blended in a Waring blender for 30 s (50 g fruit sample: 50 ml of 0.05% Na2S2O5 solution); a 2-ml aliquot was pipetted into a test tube, to which 10 ml of a binary petroleum ether/propanone mixture (97:3) was added; the tube was vortexed for 30 s, and the supernatant solution was read in a spectrophotometer at 503 and 452 nm. Lycopene and beta-carotene contents were then calculated according to the formulas indicated by Silverstein et al. (1994). For all traits, contrasts among treatments were used to quantify the effects of the background and/or the effects of heterozygous combinations at loci alc, ogc and hp. RESULTS AND DISCUSSION Total yield There were no significant differences among the genotypes with mutant genes alc, ogc and hp, indicating that heterozygosity at the alc locus, alone or in conjunction with ogc+/ogc and hp+/hp, had no effect on yield in the hybrids with a Flora-Dade x Mospomorist background (Table 1). A lack of deleterious effects of alc+/alc on yield was also found by Flori and Maluf (1994), Freitas et al. (1998), and Souza et al. (2001) in hybrid combinations, while Araújo et al. (2002) did find significant effects of these mutant genes on an inbred background.
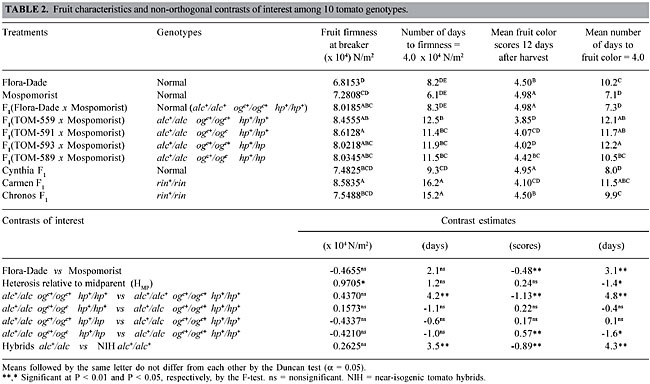
Early yield Hybrids alc+/alc, associated or not with ogc+/ogc or hp+/hp, had a tendency to produce lower early yields when compared to their NIH counterparts (alc+/alc+ ogc+/ogc+ hp+/hp+) (Table 1). This is likely a consequence of the delay in the period necessary for the fruit to reach the breaker stage brought about by the alc+/alc genotype. This tendency was not significant for hybrids with genotypic constitutions alc+/alc ogc+/ogc+ hp+/hp+, alc+/alc ogc+/ogc hp+/hp+ or alc+/alc ogc+/ogc+ hp+/hp (Table 1), but was evident in the hybrid alc+/alc ogc+/ogc hp+/hp, which had a significantly lower early yield (27.225 ton/ha) than that of the normal NIH counterpart (43.025 ton/ha). While alc+/alc reduced early yield, heterosis in the Flora-Dade x Mospomorist background increased it (+19.16 ton/ha) relative to the midparent, similar to what was found in other studies (Maluf et al., 1982; Filgueira and Leal, 1983). Mean fruit mass Background heterosis for mean fruit mass was negative (-3.5 g/fruit) and low in magnitude (-3.12%), indicating a mainly additive gene action for this trait, as found by Maluf et al. (1982) and Freitas et al. (1998). The heterozygote alc+/alc, associated or not with ogc+/ogc and/or hp+/hp, had no effect on mean fruit mass (Table 1). These results are similar to those of Leal and Mizubuti (1975), Freitas et al. (1999a) and Souza et al. (2001). However, Araújo et al. (2002) reported that mean fruit mass was significantly affected, albeit in a different genotypic background, by specific combinations at loci alc, hp and ogc; both alc/alc and alc+/alc negatively affected fruit mass. Fruit mass of alc+/alc fruit was reduced by hp/hp, but was unaffected by hp+/hp. Number of days from anthesis to fruit breaker stage Normal background (alc+/alc+) heterosis was significantly negative (-3.4 days) for this trait, suggesting partially dominant gene action in the direction of shorter time spans. Fruit of the hybrid alc+/alc ogc+/ogc+ hp+/hp+ remained on the vine before harvest two days longer than the fruit of its normal (alc+/alc+ ogc+/ogc+ hp+/hp+) NIH counterpart; even though the difference was not significant at a = 0.05, it indicates a delay in the beginning of the ripening process due to the genotypic constitution alc+/alc (Table 1). Differences among the four experimental hybrids bearing the mutant genes alc, ogc or hp were negligible, with a maximum 0.4 day difference between these genotypes (Table 1), indicating that neither ogc+/ogc nor hp+/hp had any additional effect on the trait. Fruit firmness The F1 hybrid (Flora-Dade x Mospomorist) had a significantly higher initial firmness than the midparent (Table 2); background heterosis was +13.77%, in the direction of higher firmness, an indication of some degree of dominance of the genes that control fruit firmness in a normal (alc+/alc+ ogc+/ogc+ hp+/hp+) background. At harvest (breaker stage), the NIH lines bearing genes alc, ogc and hp gave the highest firmness values, ranging from 8.02 x 104 N/m2 to 8.61 x 104 N/m2 (Table 2), but these values were not significantly different from the NIH alc+/alc+ counterpart or the hybrid Carmen F1 (rin+/rin) (8.5 x 104 N/m2), currently the standard long-shelf life tomato in the Brazilian market. Flora-Dade, an open pollinated line with an acceptable degree of firmness, gave the lowest initial values (6.81 x 104 N/m2). Differences among the hybrids became more evident as the maturation process progressed: fruit of F1(TOM-559 x Mospomorist) (alc+/alc ogc+/ogc+ hp+/hp+), F1(TOM-593 x Mospomorist) (alc+/alc ogc+/ogc+ hp+/hp), F1(TOM-589 x Mospomorist) (alc+/alc ogc+/ogc hp+/hp) and F1(TOM-591 x Mospomorist) (alc+/alc ogc+/ogc hp+/hp+) took an average of 4.2, 3.6, 3.2 and 3.1 days longer, respectively, than the normal NIH F1(Flora-Dade x Mospomorist) (alc+/alc+ ogc+/ogc+ hp+/hp+) to reach a firmness reading of 4.0 x 104 N/m2. Differences brought about by ogc+/ogc and/or hp+/hp, along with alc+/alc, were not significantly different from alc+/alc alone (Table 2). This indicates a superior shelf life of the alc+/alc hybrids relative to the normal alc+/alc+ tomatoes, as also reported by Mutschler et al. (1992), Flori and Maluf (1994), Resende et al. (1997), Freitas et al. (1998) and Souza et al. (2001), but no effects of ogc+/ogc and hp+/hp on this trait.
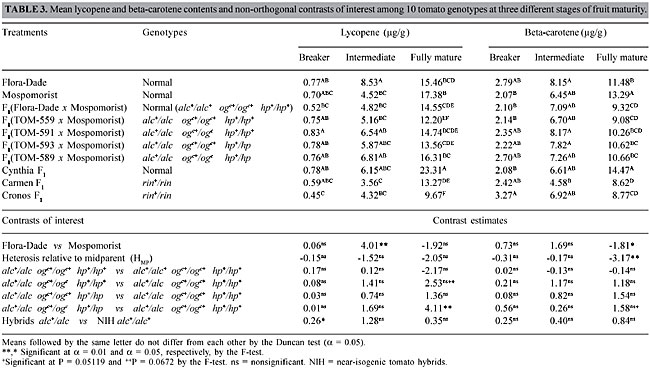
Fruit color Flora-Dade fruits took 3.1 days longer to reach fruit color score 4 than Mospomorist fruits (Table 2). Normal background (alc+/alc+) heterosis was negative, indicating dominant genetic effects in the direction of faster color development. Fruit color was affected by the presence of the mutant genes alc, ogc and hp. At the 12th d.a.h., the highest color scores were those of the NIH with the normal (alc+/alc+ ogc+/ogc+ hp+/hp+) genotype. The difference (-1.13 score units) between genotypes alc+/alc ogc+/ogc+ hp+/hp+ and alc+/alc+ ogc+/ogc+ hp+/hp+ indicates that alc+/alc had a significant effect in reducing the final fruit color score. This slower rate of fruit color development in alc+/alc relative to alc+/alc+ has also been reported by others (Piccinino and Scott, 1985; Freitas et al., 1998; Souza et al., 2001; Araújo et al., 2002). The genotype alc+/alc, in the absence of ogc+/ogc or hp+/hp, delayed the period necessary to reach fruit color score 4 by 4.8 days (Table 2). This delay was partially overcome with ogc+/ogc or hp+/hp in the alc+/alc hybrids, particularly in the genotype alc+/alc ogc+/ogc hp+/hp (Table 2), which had a significant improvement of 0.57 points in the color score 12 d.a.h. compared to alc+/alc ogc+/ogc+ hp+/hp+. The improvement brought about by the ogc+/ogc hp+/hp combination reduced the number of days necessary to reach color score 4 (Table 2). Beneficial effects of ogc+/ogc and hp+/hp on fruit color were also found by Freitas et al. (1999b) and Araújo et al. (2002). Carotenoid pigment contents There were no significant differences in lycopene or beta-carotene contents among NIH lines at the breaker and intermediate stages of maturity (Table 3). In fully mature fruit ogc+/ogc slightly improved (P = 0.0612, Table 3) the lycopene content of the alc+/alc hybrids. This improvement was reinforced by hp+/hp; alc+/alc ogc+/ogc hp+/hp had the highest lycopene content amongst all the NIH combinations (Table 3). The effects of alc, ogc and hp on beta-carotene contents were less marked than they were on lycopene. Nevertheless, alc+/alc ogc+/ogc hp+/hp had nearly significantly greater beta-carotene content than alc+/alc ogc+/ogc+ hp+/hp+ (P = 0.0519; Table 3).
CONCLUSIONS The alcobaça gene, in the heterozygous condition (alc+/alc) had no effect on total yield or mean fruit mass, but it decreased early yields, due to the increase in the period necessary for the fruit to reach the breaker stage brought about by the genotype alc+/alc. This genotype had a strong effect on fruit firmness and color as the fruit matured; alc+/alc reduced the rate of decrease in fruit firmness, and the rate of development of red fruit color. The effects of alc+/alc upon some traits were modified by ogc+/ogc and hp+/hp. Neither ogc+/ogc nor hp+/hp affected total or early yields, average fruit mass, or the number of days from anthesis to the fruit breaker stage. No improvement in fruit firmness during the fruit maturation process was brought about by ogc+/ogc or hp+/hp in alc+/alc hybrids. However, simultaneous presence of ogc+/ogc and hp+/hp in alc+/alc hybrids overcame some of the deleterious effects of the alc+/alc genotype on fruit color. External fruit color and lycopene contents were significantly improved in the alc+/alc ogc+/ogc hp+/hp hybrid, compared to alc+/alc ogc+/ogc+ hp+/hp+, and a similar tendency was found for beta-carotene. The alcobaça mutant could be effectively employed by tomato breeders for the development of tomato hybrids with extended post-harvest shelf life, especially when associated with the color enhancing mutants old gold-crimson and high-pigment in the heterozygous condition. Various of the alc+/alc hybrids studied in this experiment was competitive with at least one of the commercial hybrids used (Cynthia, Carmen or Chronos). ACKNOWLEDGMENTS The authors acknowledge the following agencies and companies in Brazil that supported this research project: CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico/MCT)/RHAE (Programa de Recursos Humanos em Áreas Estratégicas), FAEPE/UFLA (Fundação de Apoio ao Ensino, Pesquisa e Extensão/ Universidade Federal de Lavras), FAPEMIG (Fundação de Amparo à Pesquisa do Estado de Minas Gerais), CAPES/MEC, and HortiAgro Sementes. P. Moretto and V. Licursi were recipients of FINEP/CNPq scholarships. REFERENCES Araújo, M.L., Maluf, W.R., Gomes, L.A.A. and Oliveira, A.C.B. (2002). Intra and interlocus interactions between alcobaça (alc), crimson (ogc) and high pigment (hp) loci in tomato Lycopersicon esculentum Mill. Euphytica 125: 215-226. Calbo, A.G. and Calbo, M.E.R. (1989). Medição e importância do potencial da parede. Rev. Bras. Fis. Veg. 1: 41-45. Calbo, A.G. and Nery, A.A. (1995). Medida de firmeza em hortaliças pela técnica de aplanação. Hortic. Bras. 13: 14-18. Filgueira, A.V. and Leal, N.R. (1983). Avaliação dos progenitores e obtenção de novas combinações genéticas em tomate “Salada”. In: Congresso Brasileiro de Olericultura, 23. Resumos. Sociedade de Olericultura do Brasil, Rio de Janeiro, RJ, Brazil, pp. 154. Flori, J.E. and Maluf, W.R. (1994). Obtenção e avaliação de híbridos F1 de tomate (Lycopersicon esculentum Mill.) do grupo multilocular. Cienc. Prat. 18: 395-398. Freitas, J.A., Maluf, W.R., Gomes, L.A.A., Oliveira, A.C.B., Martins, W. and Braga, R.S. (1998). Padrão de amadurecimento e conservação pós-colheita de frutos de tomateiro, em função das diferentes constituições genotípicas no loco alcobaça. Rev. Bras. Fisiol. Veg. 10: 191-196. Freitas, J.A., Maluf, W.R., Azevedo, S.M., Braga, R.S. and Gomes, L.A.A. (1999a). Efeito dos alelos alc, ogc e hp e heterose em características de produção do tomateiro. Acta Sci. 21: 439-446. Freitas, J.A., Maluf, W.R., Gomes, L.A.A. and Azevedo, S.M. (1999b). Efeitos dos alelos alc, ogc e hp sobre as características de maturação e conservação pós-colheita de frutos de tomateiro. Cienc. Agrotecnol. 23: 569-577. Kopeliovitch, E., Rabinowitch, H.D., Mizrahi, Y. and Kedar, N. (1979). The potencial of ripening mutants for extending the storage life of the tomato fruit. Euphytica 28: 99-104. Leal, N.R. and Mizubuti, A. (1975). Características e conservação natural pós-colheita de frutos de híbridos entre a introdução ‘alcobaça’ e alguns cultivares de tomate. Experientiae 19: 239-257. Lobo, M. (1981). Genetic and physiological studies of the “Alcobaça” tomato ripening mutant. Ph.D. thesis, University of Florida, Gainesville, FL, USA. Lobo, M., Basset, M.J. and Hannah, L.C. (1984). Inheritance and characterization of the fruit ripening mutation in ‘alcobaca’ tomato. J. Am. Soc. HortSci. 109: 741-745. Maluf, W.R., Miranda, J.E.C. and Campos, J.R. (1982). Análise genética de um cruzamento dialélico de tomate. I- Características referentes à produção de frutos. Pesqui. Agropecu. Bras. 17: 633-634. Mutschler, M.A. (1984a). Inheritance and linkage of the ‘alcobaca’ ripening mutant tomato. J. Am. Soc. Hort. Sci. 109: 500-503. Mutschler, M.A. (1984b). Ripening and storage characteristics of the ‘alcobaca’ ripening mutant in tomato. J. Am. Soc. Hort. Sci. 109: 504-507. Mutschler, M.A., Wolfe, D.W., Cobb, E.D. and Yourstone, K.S. (1992). Tomato fruit quality and shelf life in hybrids heterozygous for the alc ripening mutant. Hort. Sci. 27: 352-355. Piccinino, L.L. and Scott, J.W. (1985). Enhancement of heterozygous nor and nor a tomato hybrid fruit color by addition of hp and/or og genes. HortSci. 20: 657 (Abstract). Resende, J.M., Chitarra, M.I., Maluf, W.R. and Chitarra, A.B. (1997). Qualidade pós-colheita em genótipos de tomate do grupo multilocular. Hortic. Bras. 15: 92-98. Silverstein, R.M., Bassler, G.C. and Morril, T.C. (1994). Identificação Espectrométrica de Compostos Orgânicos. Guanabara Dois, Rio de Janeiro, RJ, Brazil, pp. 387. Souza, J.C., Maluf, W.R., Souza-Sobrinho, F., Gomes, L.A.A., Moreto, P. and Licursi, V. (2001). Características de produção e conservação pós-colheita de frutos de tomateiros híbridos portadores do alelo “alcobaça”. Cienc. Agrotecnol. 25: 503-509. Thompson, A.E., Hepler, R.W. and Kerr, E.A. (1962). Clarification of the inheritance of high total carotenoid pigments in the tomato. J. Am. Soc. Hort. Sci. 81: 434-442. Thompson, A.E., Tomes, M.L., Erickson, H.T., Wann, E.V. and Armstrong, R.J. (1967). Inheritance of crimson fruit color in tomatoes. Proc. Am. Soc. Hort. Sci. 91: 495-504. Tigchelaar, E.C., McGlasson, W.B. and Buescher, R.W. (1978). Genetic regulation of tomato fruit ripening. Hort. Sci. 13: 508-513. Vilas Boas, E.V.B., Chitarra, A.B., Maluf, W.R. and Chitarra, M.I.F. (1999). Influência do alelo alcobaça em heterozigose sobre a vida de prateleira e qualidade pós-colheita de tomates. Cienc. Agrotecnol. 23: 650-657. |
|