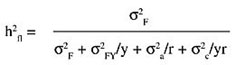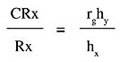![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
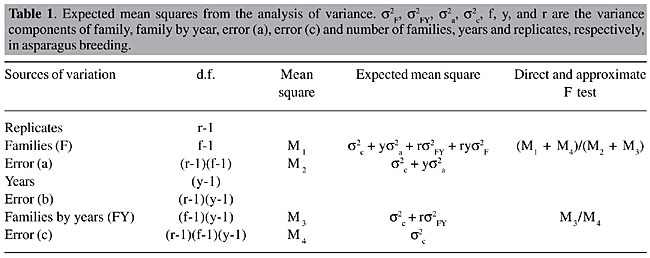
ABSTRACT. Despite the continuous breeding that has been conducted with asparagus (Asparagus officinalis L.) since the beginning of the last century, there is little information on parameters for predicting direct and indirect selection response. Yield traits for blanched asparagus production were studied along a two-year period in a half-sib family population planted in Zavalla, Argentina. Half-sib family mean heritability values were low for total yield and marketable spear number (0.31 and 0.35), intermediate for marketable yield and total spear number (0.55 and 0.64), and relatively high for spear diameter and spear weight (0.75 and 0.74). An average increase in marketable yield of 15.9% is expected after each cycle of selection of the top 5% of the families. Total yield failed to express significant genetic correlations with any of the yield components; meanwhile marketable yield showed highly significant relations with market spear number (0.96) and spear weight (0.89). Indirect selection response over yield components (CRx) failed to be advantageous over direct selection (Rx), since the ratio CRx/Rx was always equal or below unity. Key words: Asparagus officinalis, Genetic correlations, Genetic gain INTRODUCTION Asparagus (Asparagus officinalis L.) is a dioecious plant reproduced mainly by seed. Despite its perennial nature, two years of harvesting have proven to give a reliable evaluation of yield performance (Bussell et al., 1987). Different approaches in breeding programs with the aim to increase yield and uniformity have led to the release of different kinds of materials since the beginning of the last century, i.e., populations improved by mass selection, and single, double and clonal hybrids with an average yield increase of 75% over unselected materials (Ellison, 1986). Sexual dimorphism, in which staminate plants produce a higher number of thinner spears than pistillate plants, has been reported for this species (Robbins and Jones, 1926). During the last 40 years, emphasis has been given to all-male hybrids, the progeny of a supermale (YY) and a pistillate (XX) plant. However, its convenience has been argued (Corriols, 1984). Results from the Second International Asparagus Cultivar Trial (Benson, 1999) had ranked all-male hybrids on average at 14th in comparison with dioecious hybrids at the 9th position. From the selection point of view, all-male materials are dead-ends. The recombination of selected andromonoecious plants (sources of supermales) can also lead to an increased expression of andromonoecism, which restores the advantage of all-male materials (Sneep, 1953). Breeding programs require stages of selection for parents and progenies. The selection criteria of elite plants and progenies depend on the variability in the base population and the relative magnitude of the genetic components determining the phenotypic expression of the traits. Individual plant selection can be effective only if the variables under selection have high heritability values. Broad-sense heritability estimates on a one-replication basis were presented for asparagus by Legg et al. (1968) and López Anido et al. (1997). The estimates of heritability were low for most yield traits, suggesting the influence of the micro-environmental conditions upon the phenotypic expression of a single plant. Narrow-sense heritability estimates based on means of asparagus families suggested the use of recurrent selection in the breeding programs (López Anido et al., 1999). Traits under selection are often associated with each other in a very complex way. Correlations between characters have been studied to identify those of easy measurement for indirect selection for yield. Currence and Richardson (1937), Ellison et al. (1960) and Ellison (1986) found that spear diameter and spear number were highly positively correlated with yield, and negatively correlated with each other (Ellison and Scheer, 1959). Cointry et al. (2000) found that spear number and mean spear weight were the principal components of yield. All these studies were based on phenotypic correlations. For yield components to be useful as secondary traits in indirect selection, additive genetic correlations should be considered instead. Heritability and expected selection response for yield and its components would aid in decision making in the breeding programs of this species. MATERIAL AND METHODS Plant material and evaluation The open pollinated cultivar Argenteüil was a parent from which many of the modern varieties and clonal hybrids were developed (Knaflewski, 1996). Populations grown from this cultivar (cv.) are regarded as representative for the estimation of genetic parameters. In the summer of 1994 a four-year-old field of cv. Argenteüil was left for open pollination. One hundred fruits were collected from 32 plants in the fall. This constituted a base population of 32 half-sib families for our study. One-year-old crowns from these families were planted in August 1996 at the Rosario National University experimental fields located at Zavalla (33° 01’ S, 60° 53’ W), Santa Fe Province, Argentina. A randomized complete block design with four replicates of 20 plants per family was used in a normal planting grid for white asparagus (2.1 m between rows and 0.45 m among plants in the row). During the spring seasons of 1997 and 1998 the following traits of white asparagus production were evaluated on a plot basis: total spear number, mean spear weight (g), mean spear diameter (mm), measured at the base of the upper third of the spear, total yield (g), marketable yield (g) or weight of spears with diameter equal or greater than 12 mm, and number of marketable spears. All observations were conducted during a forty-day period after the harvest of the first spear of each plant. Harvests were made three times a week and spear length was standardized to 15 cm prior to weighing with a digital scale. Data analysis Data were subjected to an ANOVA using PROC GLM (SAS Institute, 1996) along the following linear model for perennial plants proposed by Nyquist (1991), but considering one location: Pjkm = µ + Rj + Fk + a(jk) + Ym + b(jm) + FY(km) + c(jkm) where Pjkm is the phenotypic plot value of the jth replicate of the kth family in the mth year, µ is the general mean, Rj is the effect of the jth replicate, Fk is the effect of the kth family, Ym is the effect of the mth year, FY(km) is the interaction effect of the kth family with the mth year, and a(jk), b(jm) and c(jkm) are error terms. The expected mean squares (MS) were deduced considering all effects randomly (Table 1). When the family by year MS was not significant, the family MS was tested against error (a) MS. Narrow sense heritability in a half-sib family mean basis was calculated as proposed by Nyquist (1991):
where s2F, s2FY, s2a and s2c are the variance components of family, family by year, error (a) and error (c) respectively, y is the number of years and r is the number of replicates. s2F represents 1/4 of additive plus 1/16 additive-by-additive variance. Confidence intervals for heritability were obtained following Knapp et al. (1985). The expected selection response (R) was estimated considering the half-sib family selection method described by Nyquist (1991): Rx = i h2fl sx where i is the selection intensity (2.06 for the top 5%), and sx is the square root of the phenotypic variance among families. Confidence intervals for the selection response were estimated as proposed by Tai (1989). The cross product sum matrix for each of the elements of the model was generated by the PROC MANOVA procedure (SAS Institute, 1996) and the genetic co-variance between traits solved. Genetic correlations were calculated as proposed by Searle (1961): rg = Covgxy/(sgx . sgy) where Covgxy is the genetic co-variance between characters x and y and sgx and sgy are the genetic standard deviations. The standard error (SE) of the genetic correlation was obtained following Becker (1967). The statistical significance of estimated genetic correlations was assessed by the test proposed by Hébert et al. (1994), in which the inferior limit of the confidence interval of genetic correlation is: rg(min) = ½rg½ - t[0.975, (f-2)] SE(rg) where t is the Student test value and f the number of families. The rg(min) value was compared to the critical absolute values of the correlation coefficients that were significant at a confidence level of 95 and 99%. For our sample size these critical values were 0.34 and 0.44, respectively (Snedecor, 1956). If the inferior limit was greater than these critical values then the genetic correlation was considered to be significantly different from zero for the corresponding confidence level. The merit of indirect selection upon yield components considering equal intensity of selection in the primary and secondary traits was tested with the ratio proposed by Falconer and Mackay (1996):
where CRx is the correlated response of character X resulting from selection applied to the secondary character Y, Rx is the direct response of selecting the desirable primary character X, hy and hx are the square roots of the heritability of characters x and y.
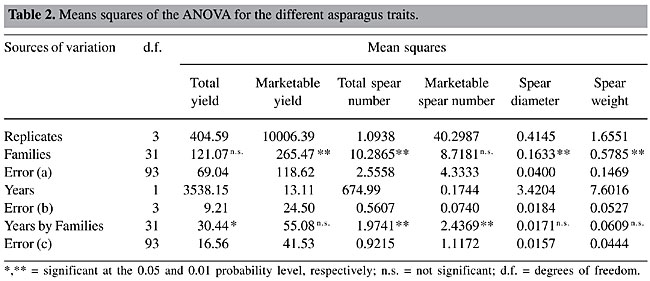
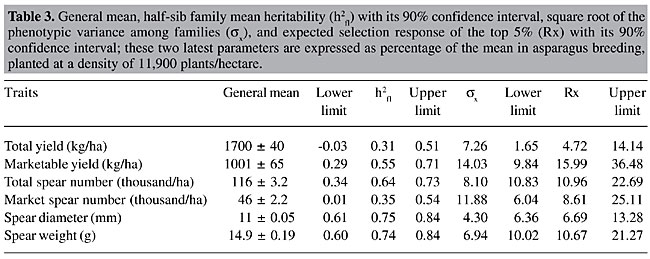
RESULTS AND DISCUSSION The ANOVA layout is presented in Table 2. Differences among families were highly significant (P < 0.01) for marketable yield, total spear number, spear diameter, and spear weight. The interaction family by year was significant (P < 0.05) for total yield and highly significant for total spear number and marketable spear number. The general mean, half-sib family mean heritability, square root of the phenotypic variance among families, and expected selection response of the top 5% were determined (Table 3). The heritability values were low for total yield and marketable spear number, intermediate for marketable yield and total spear number, and relatively high for spear diameter and spear weight.
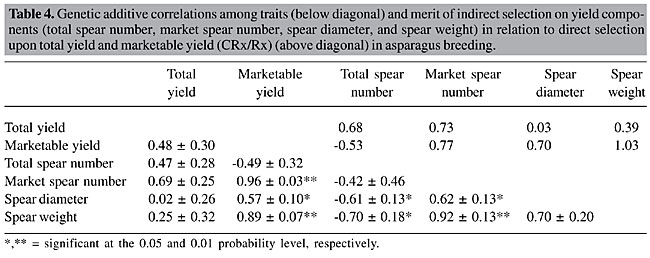
Comparing the two yield traits, total yield and marketable yield, the latter proved to be more appropriate for selection schemes, since the variation among family means was highly significant with a concomitant higher heritability value than the former. An important fraction of the variance among families was of an additive nature, thus susceptible to be used to attain selection response. From the economic point of view it would be also more appropriate to focus exclusively on marketable yield, since even when the rejected fresh marketable production is sold for processing industry, it demands an increased labor and handling cost per harvested kg (Van den Broek and Boonen, 1990). Spear diameter and spear weight gave the highest heritability estimates. This is in concordance with the close relation between spear diameter of parental plants and offsprings found by Currence (1947) in cv. ‘Mary Washington’. High heritabilities for marketable yield and spear diameter (0.72 and 0.60, respectively) in a full-sib family basis were also found by López Anido et al. (1999) in a study of cv. ‘Argenteüil’ under green and blanched production. Based on our results, after each cycle of selection of the top 5% of the families an average increase of 15.9% in marketable yield is expected. Due to the higher square root of the phenotypic variance among families, the selection response was higher for marketable yield than for the rest of the traits. If we consider a five-year-selection cycle, three years from seed to end of the second harvest plus two more years for recombination and conformation of the next generation, the annual increase would be on the order of 3.2%. Analyzing the genealogy of modern asparagus cultivars (Knaflewski, 1996), we can observe that only three or at most four cycles of selection and recombination were made. If continuous recurrent family selection schemes had been maintained since 1919, when the first improved cultivar ‘Mary Washington’ was released by Norton (1919), the average yield of actual asparagus breeding stocks would be much greater. The genetic correlations and the merit of indirect selection upon yield components were calculated (Table 4). Total yield failed to express significant associations with any of the characters; meanwhile marketable yield showed a highly significant relation with market spear number and spear weight. When studying the correlations among yield components, we found that total spear number was significantly negatively associated with spear diameter and spear weight; while marketable spear number was positively associated with these two components. Thus, if selection were conducted on marketable yield, contrary to what can be inferred when considering phenotypic correlations (Ellison and Scheer, 1959), it would be possible to attain increases in both marketable spear number and spear diameter. Indirect selection failed to be advantageous over direct selection, since the ratio CRx/Rx was always equal or below unity (Table 4).
CONCLUSIONS Substantial additive variation for marketable yield is available to be exploited by family mean selection schemes. When selecting for this trait, no negative effect on marketable spear number or spear diameter will be produced. Indirect selection for yield components is not advantageous over direct selection. REFERENCES Becker, W.A. (1967). Manual of Procedures in Quantitative Genetics. Washington State University Press, Pullman, WA, USA. Benson, B.L. (1999). Second international asparagus cultivar trial. Acta Hortic. 479: 143-148. Bussell, W.T., Falloon, P.G. and Nikoloff, A.S. (1987). Evaluation of asparagus yield performance after two years’ harvesting. N.Z.J. Exp. Agric. 15: 205-208. Cointry, E., López Anido, F.S., Gatti, I., Cravero, V.P., Firpo, I.T. and García, S.M. (2000). Early selection of elite plants in asparagus. Bragantia 59: 21-26. Corriols, L. (1984). Are all-male hybrids attractive? Asparagus Res. Newsl. 2: 16-19. Currence, T.M. (1947). Progeny tests of asparagus plants. J. Agric. Res. 74: 65-76. Currence, T.M. and Richardson, A.L. (1937). Asparagus breeding studies. Proc. Am. Soc. Hortic. Sci. 35: 554-557. Ellison, J.H. (1986). Asparagus breeding. In: Breeding Vegetables Crops (Bassett, M.J., ed.). AVI Publishing Company, Westport, CT, USA, pp. 521-569. Ellison, J.H. and Scheer, D.F. (1959). Yield related to brush vigor in asparagus. Proc. Am. Soc. Hortic. Sci. 73: 339-344. Ellison, J.H., Scheer, D.F. and Wagner, J.J. (1960). Asparagus yield as related to plant vigor, earliness and sex. Proc. Am. Soc. Hortic. Sci. 75: 411-415. Falconer, D.S. and Mackay, T.F.C. (1996). Introduction to Quantitative Genetics. 4th edn. Longman, Essex, England. Hébert, D., Fauré, S. and Olivieri, I. (1994). Genetic, phenotypic, and environmental correlations in blck medic, Medicago lupulina L., grown in three environments. Theor. Appl. Genet. 88: 604-613. Knaflewski, M. (1996). Genealogy of asparagus cultivars. Acta Hortic. 415: 87-91. Knapp, S.J., Stroup, W.W. and Ross, V.M. (1985). Exact confidence intervals for heritability on a progeny mean basis. Crop Sci. 25: 192-194. Legg, P.D., Souther, R. and Takatori, F.H. (1968). Estimates of heritability in Asparagus officinalis from replicated clonal material. Proc. Am. Soc. Hortic. Sci. 92: 410-417. López Anido, F.S., Cointry, E.L., Picardi, L. and Camadro, E. (1997). Genetic variability of productive and vegetative characters in Asparagus officinalis L. - Estimates of heritability and genetic correlations. Braz. J. Genet. 20: 275-281. López Anido, F.S., Cointry, E.L., Firpo, I.T. and García, S.M. (1999). Heritability of green and white asparagus yield. In: Eucarpia Leafy Vegetables’ 99 (Lebeda, A. and Kristková, E., eds.). Palacký University Press, Olomouc, Czech Republic, pp. 277-280. Norton, J.B. (1919). Washington asparagus: Information and suggestions for growers of new pedigreed rust-resistance strains. U.S. Dep. Agric. Bur. Plant Ind. Circ. 7. Nyquist, W.E. (1991). Estimation of heritability and prediction of selection response in plant populations. Crit. Rev. Plant Sci. 10: 235-322. Robbins, W.W. and Jones, H.A. (1926). Sex as a factor in growing asparagus. Proc. Am. Soc. Hortic. Sci. 25: 13-16. SAS Institute (1996). SAS User’s Guide: Statistics. SAS Inst. Inc., Cary, NC, USA. Searle, S.R. (1961). Phenotypic, genotypic and environmental correlations. Biometrics 17: 474-480. Snedecor, G.W. (1956). Statistical Methods. 4th edn. Iowa State University Press, Ames, IA, USA. Sneep, J. (1953). The significance of andromonoecy for the breeding of Asparagus officinalis L. II. Euphytica 2: 224-228. Tai, G.C.C. (1989). A new procedure to construct confidence intervals for genotypic variance and expected response to selection. Genome 32: 307-308. Van den Broek, J.H. and Boonen, P.H. (1990). Today’s asparagus breeding in the Netherlands. Acta Hortic. 271: 33-38. |
|