![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
ABSTRACT. South America is responsible for about half of the cassava world production. In the 1970’s productivity of the crop on the continent was about 15 ton/ha, and dropped continuously until reaching 12 ton/ha in 2004. India’s productivity of cassava increased from 10 ton/ha in the 1970’s to 28 ton/ha in 2004. Brazil contributed significantly to improving cassava crops through the Instituto Agronômico de Campinas in the 1960’s and 1970’s. The Universidade de Brasília released high-protein content hybrids, apomictic clones and explored the potential of indigenous landraces. Key words: Cassava, India, Plant breeding, Productivity INTRODUCTION Cassava is an important subsistence crop in South America (FAO, 2004). Because of its high calorie productivity (Coursey and Haynes,1970), biological efficiency as an energy producer (Vries et al., 1967), year round availability, tolerance to extreme stress conditions, adaptations to a wide range of ecological conditions (Jones, 1970), and its ability to grow in suboptimal soil (Papadakis, 1966), cassava can play a major role in alleviating the continent’s food supply problem. In the early 1970’s the U.S. president, upon being alerted to developing countries’ food crisis, appointed a Science Advisory Committee Panel on the world’s food supply to report on food research priorities. It recommended that the agricultural potential of vast areas of uncultivated land, particularly in South America and Africa, be thoroughly evaluated for sustained food production and placed emphasis on cassava as a selected crop which has the potential for satisfying the huge demand for food (Hendershott, 1972). Since that time, much attention has been paid to cassava as a priority crop in the newly created International Centers for Agricultural Research. The International Center for Tropical Agriculture, at Cali, Colombia, was given a mandate for improving cassava throughout the world and in South America in particular. PRODUCTIVITY DURING THE LAST 20 YEARS Productivity per hectare is the criterion for assessing crop improvement. In this context, on the basis of FAO data, and the statistical yearbooks of the same organization over the last 30 years, the following can be noted:
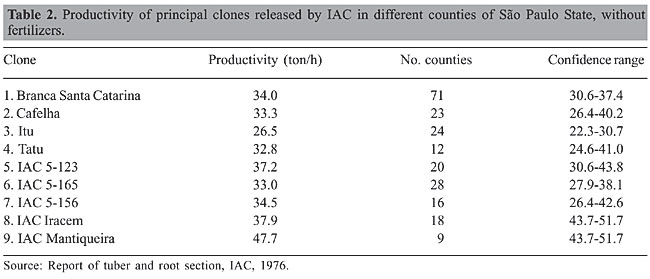
CAUSES OF DETERIORATION IN CASSAVA PRODUCTIVITY The deterioration in cassava productivity on the continent during the 1970’s and the 1980’s is due to the following facts. The principal producer of the continent, São Paulo State, contributed about 1/3 of the total production of the country with an average productivity of 21 ton/ha. This level of productivity was possible because of the dedicated, brilliant and conscientious work of the Instituto Agronômico de Campinas (IAC) and its cassava breeders (Table 2). Since the beginning of the 1970’s, the farmers of São Paulo replaced cassava cultivation with other cash crops. The cassava area decreased in the country and the whole continent. In the meantime, cassava cultivars planted in other Brazilian states did not show the good performance of IAC’s cultivars in São Paulo State. Consequently, this contributed to reducing cassava productivity in Brazil and the whole continent.
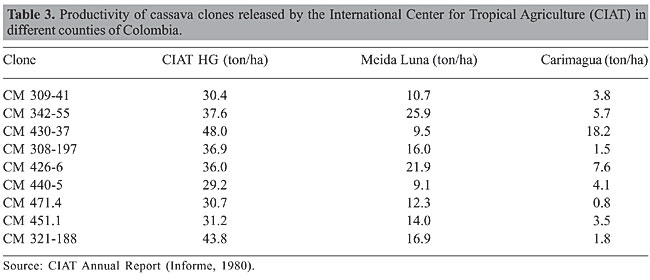
THE SUCCESS OF INSTITUTO AGRONÔMICO DE CAMPINAS CULTIVARS The highly productive, well-adapted IAC cultivars are simple products of the cassava breeding program in this institution. The IAC cassava program began by determining good progenitors from whose progeny new cultivars could be selected on the basis of productivity and resistance to diseases and insects. This was carried out through comprehensive tests of combining ability, which included clones collected from São Paulo and Minas Gerais States. In these states, wild species of Manihot normally grow very close to cultivated cassava clones and natural interspecific hybridization frequently occurs between them (IITA, 1983; Nassar, 1984). Progeny seedlings of these natural crosses grow simultaneously, and some of them are selected by farmers, reproduced vegetatively, giving rise to new clones. These clones grow in commercial plantations and are subject to auto-pollination due to the monoclonal system of plantations (Nassar and O’Hair, 1985). Emerging homozygous plants have genes of wild species introgressed into their genomes. This cycle of hybridization is repeated in nature and inbred clones enriched by high adaptive genes of the wild species come to cultivations by farmers. These clones are the type of clones collected by IAC breeders and used in their combining ability trials. Among them were the most successful clones ever known in the history of cassava: Branca Santa Catarina, Mantiqueira, Engana Ladrão, and others. In its evaluation, to release new cultivars, the IAC carried out performance trials of root yield under suboptimal conditions of soil and mineral nutrients to ensure the largest amplitude of cultivar adaptations. Through a number of counties, ranging from 9 to 70, these cultivars showed the highest productivity compared to those released by other institutions (Tables 2 and 3).
The combining ability approach followed by the IAC to produce superior progeny and perpetuate selected individuals by cloning showed more potential and effectiveness than the method of recurrent selection followed by other institutions. It seems that nonadditive genes and heterosis are predominant in cassava as modes of gene action. PROSPECTS OF IMPROVEMENT IN THE NEAR FUTURE The University of Brasília adopted three lines of research which has been carried out by this researcher since 1975. Exploration of wild-genetic resources Cassava originated in the Northern Amazon, South America (Nassar, 1978a). In various parts of the continent, wild relatives grow naturally and exhibit a vast array of genetic variations yet to be explored and utilized as a source of many genes (Nassar, 1978b; Nassar, 1986). In the meantime, many leading scientists emphasized the valuable sources of wild species in breeding crops (Monaco and Carvalho, 1971; Frey, 1976; Rick, 1978). However, very little work has been done to systematically improve cassava by introgressed genes from wild species up to the mid 1970s. Beginning at that time, wild species from the whole continent were collected, evaluated and maintained in a living collection. Interspecific hybridizations were carried out systematically by Pereira et al. (1981). Interspecific hybrids were produced and used in the program at Brasília, while some of them were donated to IITA in 1979, 1980, 1981. These hybrids and their progenies were developed by Hahn et al. (1990), an IITA breeder, and gave rise to the most resistant cultivars to mosaic ever known in West Africa. The cultivars of the family MS raised from these hybrids cover now 4 million hectares in Nigeria and brought the country to be the first ranking producer all over the world (Nassar and Grattapaglia, 1986). The following is a summary of breeding trials with these species:
Production of polyploid hybrids by the manipulation of meiotic restitution Tetraploid and triploid cultivars of vegetatively reproduced crops have been successfully used by plant breeders. Trials of chromosome doubling in cassava were made early in the 1960’s by using colchicines. However, this attempt has not led to the development of tetraploid or triploid cultivars, probably due to instability of the chimera produced. Recently, this author (Nassar, 2004) has discovered that interspecific hybrids of cassava exhibit a meiotic irregularity accompanied by high frequency of meiotic restitution which leads to the production of 2n hybrid diploid gametes. The author manipulated this phenomenon to produce a productive triploid that was tolerant to drought conditions (Nassar, 1991). In addition to this triploid, the author proceeded to the production of a trisomic hybrid adapted to savanna conditions. Interspecific hybrids of cassava with M. glaziovii, M. pseudoglaziovii, M. anomala, and M. aesculifolia were polyploidized artificially by colchicine treatment. Polyploid types of these interspecific hybrids had their fertility restored which enabled further backcrosses and manipulation towards the production of triploids. Development of apomictic true breeding lines Since cassava is vegetatively propagated by means of stem cuttings, it is considered to be a labor intensive crop. Vegetative cuttings are also often responsible for the spread of diseases and pests throughout the tropics. This author (Nassar, 1985) and O’Hair (1985) proposed the idea that the use of true seed in place of stem cuttings for cassava production would eliminate these problems and potentially reduce production costs. One limiting factor, though, is the lack of quick and uniform seed germination. Another difficulty is the genetic segregation and lack of true breeding lines. If apomictic easily germinated lines could be developed, this idea would be successful. Since 1993, this author (Nassar et al., 2000) has been involved in the use of mass selection to gradually modify the cassava population with respect to seed dormancy and has obtained an easily germinated seed population. During the last years, genes of apomixis were successfully introduced from the wild-species M. glaziovii and M. neusana. Various apomictic clones were developed by this author. Molecular markers were used to confirm the apomictic nature (Nassar et al., 2000; Nassar, 2001, 2002). EXPLORING THE POTENTIALITY OF INDIGENOUS CASSAVA CLONES Brazil is the origin of the crop where it has been domesticated by indigenous groups. Genetic diversity of its landraces is immense and yet to be explored. In the latest years, this researcher collected these indigenous clones systematically and could select from them high B-carotene content that reached 8 mg/kg combined with excellent palatability (Nassar et al., 2005). It was also possible to select some clones that are rich in lycopene having 6-8 mg/kg (see photo).
A LESSON TO BE LEARNED FROM INDIA The tripled productivity of cassava from 10 ton/ha in the 1970’s to 28 ton/ha in India has attracted scientists from all over the world. Enquiries about this magic had been always raised. Some explanations were given by Nair (2006) which are summarized here. These are:
The principal cause of this excellence is undoubtedly the leading work of CTCRI and its devoted breeders. ACKNOWLEDGMENTS Research partially supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). The above mentioned living collection of wild cassava has been established with the help of the International Development Research Center, IDRC, Ottawa. REFERENCES CIAT (1980). Annual Report. CIAT, Cali. Coursey DG and Haynes PH (1970). Root crops and their potential as food in the tropics. World Crops 22: 261-265. FAO (1989). Yearbook. FAO, Rome. FAO (2004). Production yearbook. FAO, Rome. Frey KJ (1976). Plant breeding in the seventies: useful genes from wild plants species. Egypt. J. Genet. Cytol. 5: 460-482. Hahn SK, Bai KV and Asiedu R (1990). Tetraploid, triploids, and 2n pollen from diploid interspecific crosses with cassava. Theor. Appl. Genet. 79: 433-439. Hendershott CH (1972). In preface to a literature review and research recommendations on cassava. Georgia University Press, Athens. IAC (1976). Relatório anuário. IAC, Campinas. IITA (1983). Annual report. IITA, Ibadan. Jones WO (1970). Manioc in Africa. Stanford University Press, Stanford. Monaco LC and Carvalho A (1971). Transfer of rust resistance from Coffea canifora to C. Arabica. Cienc. Cult. 23: 101-102. Nair SG (2006). Overcoming the challenge. Proceedings of the 1st Cassava Plant Breeding Meeting Biotechnology and Ecology, Brasília. Nassar NMA (1978a). Conservation of the genetic resources of cassava, Manihot esculenta Crantz: determination of wild species localities with emphasis on probable origin. Econ. Bot. 32: 311-320. Nassar NMA (1978b). Hydrocyanic acid content in some wild Manihot (cassava) species. Can. J. Plant Sci. 58: 577-578. Nassar NMA (1980). Attempts to hybridize wild Manihot species with cassava. Econ. Bot. 34: 13-15. Nassar NMA (1984). Natural hybrids between Manihot reptans Pax and M. alutacea Rogers & Appan. Can. J. Plant Sci. 64: 423-425. Nassar NMA (1985). Manihot neusana Nassar: a new species native to Paraná, Brazil. Can. J. Plant Sci. 65: 1097-1100. Nassar NMA (1986). Genetic variation of wild Manihot species native to Brazil and its potential for cassava improvement. Field Crops Res. 13: 177-184. Nassar NMA (1989). Broadening the genetic base of cassava, Manihot esculenta Crantz by interspecific hybridization. Can. J. Plant Sci. 69: 1071-1073. Nassar NMA (1991). Production of triploid cassava, Manihot esculenta Crantz. Field Crops Res. 13: 173-182. Nassar NMA (2001). The nature of apomixis in cassava. Hereditas 134: 185-187. Nassar NMA (2002). Apomixis and cassava. Genet. Mol. Res. 2: 126-128. Nassar NMA (2004). Polyploidy, chimera and fertility of interspecific cassava interspecific hybrids. Ind. J. Genet. 64: 132-133. Nassar NMA and Dorea G (1982). Protein content of cassava cultivars and its hybrid with wild Manihot species. Turrialba 32: 429-432. Nassar NMA and O’ Hair (1985). Variation among cassava clones in relation to seed germination. Indian J. Genet. Plant Breed. 45: 394-398. Nassar NMA and Grattapaglia D (1986). Variabilidade de clones da mandioca em relação à fertilidade e aspectos morfológicos. Turrialba 36: 555-559. Nassar NMA, Santos E and David SR (2000). The transference of Apomixis genes from Manihot neusana to cassava, M. esculenta Crantz. Hereditas 132: 167-170. Nassar NMA, Vizzotto CS, Schvartz CA and da Silva H (2005). Potentiality of cassava as a source of Carotenoides. J. Food Agric. Environ. 3: 33-35. Nichols RFW (1947). Breeding cassava for virus resistance. East Afr. Agric. J. 12: 184-194. Papadakis J (1966). Crop ecology survey in West Africa (Liberia, Ivory Coast, Ghana, Togo, Dahomy, Nigeria). FAO, Rome. Pereira AS, Lorenzi JO, Monteiro DA and Veiga AA (1981). Estudo de uma progenia de autofecundação de mandioca Guaxupé. Bragantia 40: 217-219. Rick CM (1978). Potential improvement of tomatoes by controlled introgression of genes from wild species. Pudoc, Wageningen, 167-173. Vries CA, Ferweds JD and Flach M (1967). Choise of crops in relation to actual and potential production in tropics. Neth. J. Agr. Sci. 15: 241-246. |
|